РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ
ЗООИНЖЕНЕРНЫЙ ФАКУЛЬТЕТ
______________________________________________________________________
ДИПЛОМНАЯ РАБОТА - УСТАРЕЛА. СМОТРИТЕ НОВУЮ РЕДАКЦИЮ
http://orlik62.narod.ru/links_diplom.html
На тему: “Анализ соответствия традиционных технологий быстроаллюрного коневодства России современным зоотехническим представлениям”
Выполнил студент 51 группы
вечернего обучения зоотехнического
факультета Дёмин И.И.
Руководитель – к.с/х.н. Парфенов В.А.
Москва – 2005
СОДЕРЖАНИЕ
Введение
Глава 1. Влияние экологических факторов на воспроизводительную функцию племенных быстроаллюрных лошадей.
Глава 2. Технологии выращивания и содержания быстроаллюрных лошадей.
2.1. Влияние условий среды и физической нагрузки на организм животных
2.2. Технология выращивания жеребят быстроаллюрных пород
2.3. Эксплуатация, тренинг быстроаллюрной лошади и организация моциона племенных быстроаллюрных лошадей
Глава 3. Организация кормления лошадей
3.1. Организация обеспечения всех половозрастных групп лошадей полноценными сбалансированными физиологически обоснованными кормами
3.2. Значение минеральных веществ в кормлении лошадей
3.3. Значение витаминов в кормлении лошадей.
3.4. Протеиновая питательность кормов и баланс аминокислот. Значение баланса аминокислот для различных половозрастных групп быстроаллюрных лошадей.
3.5. Углеводная питательность корма и значение биологических экстрактивных веществ
3.6. Липидная питательность кормов и значение жиров в жизни быстроаллюрных лошадей
3.7. Энергетическая ценность кормов
3.8. Характеристика основных кормов
3.8.1. Сено и солома
3.8.2 Перспектива применения сенажа
3.8.3. Корне- и клубнеплоды. Фрукты.
3.8.4. Веточный корм.
3.8.5. Злаковые и бобовые корма.
3.8.6. Комбикорма неполнорационные, полнорационные и условно полнорационные.
3.9. Организация прочной кормовой базы
3.10. Анализ сбалансированности по питательным веществам традиционных рационов быстроаллюрных лошадей
.
3.11. Разработка подкормки для жеребят-сосунов рысистых пород лошадей
Глава 4. Механизация производственных процессов на коневодческих предприятиях
4.1. Обеспечение комфортных условий содержания и тренинга быстроаллюрных лошадей
Глава 5. Профилактические ветеринарные мероприятия и зоогигиена
5.1. Обеспечение благополучной ветеринарно-санитарной обстановки
5.2. Значение борьбы с заразными инфекционными болезнями.
5.2.1. Вирусный артериит.
5.3. Значение противопаразитарных мероприятий
5.4. Зоогигиена и профилактика
Глава 6. Организационно-правовые мероприятия организации коневодческого дела
6.1. Рациональная расстановка, воспитание и подготовка кадров коневодства
6.2. Создание материальной и моральной заинтересованности в продолжительной и добросовестной работе.
6.3. Племенное дело
6.3.1. Значение достоверности информации используемой в племенной работе.
6.3.2. Оценка интерьера. Система модельных характеристик интерьера быстроаллюрной лошади.
6.3.3. Эффект гетерозиса и спортивное коневодство.
6.4. Организация спортивного коневодства.
6.5. Зоотехнический аудит и лицензирование в конном деле.
6.5.1. О необходимости введения в практику животноводческой отрасли зоотехнического аудита.
6.5.2. Зоотехнический аудит – организационно-экономический инструмент управления животноводческой отраслью.
6.5.3. Общая модель зоотехнического аудита
6.5.4. Общие принципы аудита систем экологического управления и менеджмента.
6.5.5. Алгоритм управления зоотехнической аудиторской деятельностью.
6.5.6. Направление применения результатов зоотехнического аудита
6.5.7. Применение зоотехнического аудита в быстроаллюрном коневодстве.
Заключение
Приложения
Литература
Введение
Основные приемы зоотехнии начали зарождаться еще при первобытнообщинном строе, одновременно с возникновением животноводства. За всю историю доместикации человек предъявлял животным различные требования, которые произрастали из его собственных потребностей. Животноводство служит человеку источником высокоценных продуктов питания (молоко, мясо, яйцо и т.п.), сырья для промышленности (шерсть, мех, кожевенное сырье, пух и т.п.) и энергии для вспашки, перевозки грузов и т.п.
На первом этапе человек был озадачен выбором животных с полезными для него отклонениями признаков, закреплением этих выбранных отклонений в потомстве путем отбора в производящий состав лучших животных.
В последствие в зоотехническую культуру влилось понятие подбора, которое вместе с отбором и подразумевают целенаправленную деятельность человека по совершенствованию наследственного потенциала полезных признаков сельскохозяйственных животных. Отбор и подбор и по ныне являются сутью селекции.
Накопив определенный потенциал знаний, человек сформулировал для себя ещё более широкое понятие – племенная работа. Значение нововведения состояло в том, что наследственные задатки хозяйственно-полезных признаков животные способны проявлять только в определенных условиях внешней среды, названных человеком – комфортными. С тех пор в понятие “племенная работа” вкладывается смысл не только “собственно селекция”, но и обеспечение этой работы комфортными условиями внешней среды (1), а также физиологически обоснованной организацией кормления, при ведении селекции в популяции здоровых животных с обязательным соблюдением санитарно-ветеринарных требований к условиям содержания.
Однако даже при полном понимании значимости каждой составной части, племенное дело было поставлено человеком себе на службу не одинаково в различных отраслях животноводства. К настоящему моменту наибольшего расцвета племенная работа достигла в птицеводстве, аквакультуре и свиноводстве, а наименьшего, по нашему мнению, в овцеводстве и коневодстве.
На протяжении уже 6 тысяч лет лошадь является верным спутником человека. На протяжении этого времени роль лошади в социально-экономической сфере человеческой цивилизации изменялась в соответствии с изменениями уклада жизни человека (2). Значение лошади в жизни человека обеспечивалось универсальностью её использования, высокая интенсивность роста молодняка, низкая себестоимость работ и продукции, высокая биологическая ценность конины и молока.
В эпоху научно-технического прогресса машины заменили лошадей во многих областях, тем не менее, интерес к этим животным не ослабевает до сих пор.
В течение нескольких десятилетий мировое поголовье лошадей зафиксировалось на уровне 65 миллионов голов (3). По мере того, как меняется жизнь человека, изменяется и интерес человека к лошади. Наиболее динамично лошадь “завоевывает сферы спорта, развлечений, активного отдыха, восстановления здоровья людей, производства уникальных продуктов питания” (3). Ипподромный бизнес, как самостоятельная отрасль, но опирающаяся на скаковое и рысистое коневодства, достиг денежного оборота в десятки миллиардов долларов ежегодно.
Однако, несмотря на структурную перестройку коневодства и высокую динамику освоения новых сфер деятельности лошадьми, технологии разработанные человеком в период от начала доместикации до момента создания чистокровной верховой породы, к настоящему времени мало изменились.
В птицеводстве, аквакультуре и свиноводстве в полном объеме пользуются преимуществами комфортных условий содержания животных, широко применяется промышленное скрещивание. В тоже время в коневодстве этим приемам зоотехнии уделяется недостаточное внимание. Усугубляется это обстоятельство ещё и тем, что северные и центральные регионы России с их климатическими условиями, относятся к зонам рискового животноводства, по причине значительного влияния внешней среды на хозяйственно-полезные качества выращиваемых животных на пастбищах и в табунах.
Обзорно-аналитический характер данной дипломной работы позволяет сопоставить соответствие традиционных технологий в быстроаллюрном коневодстве России научным знаниям в области зоотехнии на современном этапе.
В данной дипломной работе в качестве практического приложения приводится разработка сбалансированной подкормки жеребят-сосунов рысистых пород лошадей. Также приводится разработка концептуальных основ зоотехнического аудита.
Глава 1. Влияние экологических факторов на воспроизводительную функцию племенных быстроаллюрных лошадей.
Основное условие, определяющее динамику развития коневодческой отрасли – организация расширенного воспроизводства конского поголовья (2).
Длительное время доминировал подход к изучению влияния факторов внешней среды на лошадей в совокупности со стремлением максимального снижения себестоимости выращиваемых лошадей. По-нашему мнению, данный подход может широко применяться для продуктивного и рабочепользовательского коневодств, основанных на экстенсивных табунно-пастбищных технологиях и совершенно недопустимых для быстроаллюрного коневодства.
Воспроизводство лошадей складывается в основном из пяти основных периодов – оплодотворения кобыл, их жеребости, выжеребки, выращивания молодняка и выдержки племенной быстроаллюрной лошади. Все эти стадии требуют глубоких знаний физиологии содержания, тренинга, кормления и ухода за лошадьми, умения грамотного применения этих знаний для формирования эффективных технологических процессов на основе современных представлений о приемах и способах животноводства.
В комфортных условиях содержания, полноценного и сбалансированного кормления, реализуются заложенные в зиготе наследственные свойства родительских пар. В таких условиях влияние внешней среды на организм животного минимальный и процессы роста и развития формируют организм животного максимально соответствующий его геному.
Содержанию и кормлению кобыл в период беременности человек старается уделять большое внимание. Однако очень редко человек уделяет внимание экологии и не только содержанию в корме, кормовых добавках и воде загрязняющих веществ (нитраты, удобрения не биологического происхождения, ядохимикаты, гербициды, тяжелые металлы, нефтепродукты, консерванты, красители и т.д.), но и содержанию вредных веществ в окружающей жеребую лошадь среде.
При действии на организм животного неблагоприятных факторов, проявляется морфо-физиологическая адаптация, проявляющаяся в координации приспособительных процессов при помощи двух главных систем – нервной и гуморальной.
Общеизвестно, что любое живое существо, приспосабливается к условиям своего обитания: сезонные явления, температурный режим, влажность, давление, солнечная радиация, состав пищи, воды, воздуха и др. При воздействии на организм животного извне любых химических веществ возникает ответ – синтез соответствующих гормонов и ферментов, необходимых для обеспечения усвоения, путем трансформации в доступные для потребления организмом формы. Приспособление к химизму и характеру внешних условий отражается в устройстве систем потребления и обмена с внешней средой (пищеварение,
дыхания, терморегуляции, выделения и т.п.), а также выражается в иммунных реакциях организма и составе иммунного фона. При попадании в организм нового вещества, не появлявшегося ранее, начинается синтез новых форм ферментов, а также изменяются гормональный фон и иммунные реакции. Таким образом, любые качественные и количественные изменения состава веществ, поступающих из внешней среды, неминуемо приводят к генетически обусловленного состава ферментов, гормонального и иммунного фонов организма.
Из эмбриологии и иммунологии известно, что между организмом матери и плодом существует иммунологическая несовместимость. Плод, имеющий геном, отличный от материнского генома (на 50%), является для неё чужеродным. По этой причине, между ними всегда существует иммунный конфликт, подавляемый механизмом, способствующим сохранению плода и препятствующим его отторжению. Плацента и плод синтезируют ряд белковых факторов и гормонов (эстрогены, простагландины, прогестерон), подавляющие реакцию отторжения (6).
При попадании в организм жеребой кобылы новых веществ или их нового количества, её организм адаптируется к новым веществам, а вот плод этой возможности практически не имеет. В результате биохимической адаптации кобылы может возникнуть тканевая несовместимость с неадаптированным плодом. В результате иммунологической атаки на плод нарушаются условия его питания, роста и развития, вследствие чего могут возникнуть отклонения в его развитии, уродство и аллергия(6).
Для убедительности и конкретизации вышеизложенного приведем ниже информацию о некоторых проведенных исследованиях.
Так было получено значительное количество подтверждающих данных о том, что замедление развития плода в матке является очень важным фактором риска возникновения сердечно-сосудистых нарушений и нарушения обмена веществ во взрослом возрасте. Эксперименты же, проводимые на животных (морские свинки, крысы, овцы), подтвердили взаимосвязь между задержкой внутриутробного развития и формированием артериальной гипертензии или обменных нарушений во взрослом возрасте. Независимо от причины, приводящей к нарушению внутриутробного развития у животных (нарушение маточного кровотока, анемия у матери, недостаточное калорийное питание, либо низкобелковая диета во время беременности), результат получался один и тот же (6).
|
|
|
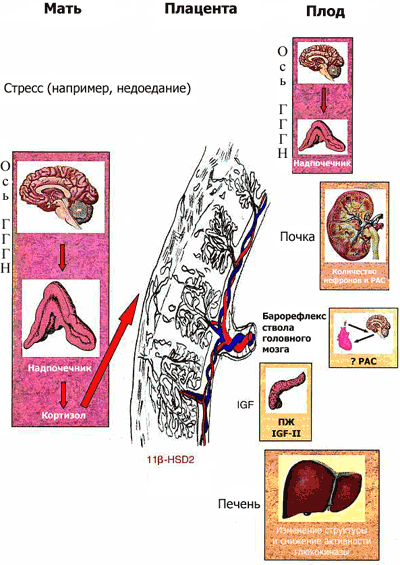
Рис. 1 Потенциальные механизмы "программирования плода"
Стресс материнского организма (например, недоедание во время беременности) приводит к активации оси гиппокамп-гипоталамус-гипофиз-надпочечники (ГГГН) у беременной, что, в свою очередь, в зависимости от уровня активности фермента плаценты, инактивирующего кортикостероиды (11 -гидроксистероиддегидрогеназы 2-го типа, 11 HSD-2), приводит к воздействию на плод высоких концентраций глюкокортикоидов (ГК). Высокий уровень глюкокортикоидов в крови плода способствует изменению числа стероидных рецепторов в ГГГН системе, изменяет уровень активности других функциональных систем (гормон роста /инсулин-зависимый фактор роста, ренин-ангиотензиновая система, вызывает замещение - клеток поджелудочной железы плода - клетками, характерными для взрослого организма), что в конечном итоге приводит к патологическим изменениям во взрослом возрасте. IGF II: инсулиноподобный фактор роста II; РАС: ренин-ангиотензиновая система; ПЖ: поджелудочная железа (6). |
Потомство этих животных имело:
- существенные изменения в морфометрии печени и значительное снижение активности глюкокиназы в перивенозных областях (у крыс);
- артериальную гипертензию, уменьшение объема мышечной ткани и увеличение жировой ткани, нарушение толерантности к глюкозе, нарушенный гомеостаз холестерина (у морских свинок);
- артериальную гипертонию к 3-х месячному возрасту (у овец) (6).
Имеются также неопровержимые данные о том, что реализация генетической программы формирования головного мозга осуществляется во взаимодействии со средовыми факторами. Эмбриональный и ранний постэмбриональный периоды развития животного играют исключительно важную роль в формировании головного мозга
.
Рост и развитие мозга может нарушаться под влиянием ряда факторов:
- высокой чувствительностью мозга в критические периоды развития к внешним воздействиям,
- последствиями воздействий (нейро-гуморальные, токсические, физические, медикаментозные и т.п.), имеющими необратимый характер, как постоянно действующие факторы.
Необратимость последствий воздействий зависит, прежде всего, от того, что зрелые нервные клетки (нейроны) не обладают способностью к делению, в связи, с чем любые факторы, которые приводят к уменьшению их численности к моменту рождения, делают мозг малонейронным, вследствие, уменьшения интенсивности деления нейробластов или увеличение числа погибающих нейробластов и нейронов, с автоматическим отклонением от нормального процессов формирования связей между нейронами - образования синапсов (7).
В исследуемых литературных данных указано на то, что у разных линий крыс животные с большей массой мозга обладают лучшими способностями к обучению. Мыши с большей массой головного мозга быстрее достигают критерия обученности и успешнее обучаются в дальнейшем (7).
Также обнаружена и закономерность, что факторы, приводящие к уменьшенной массе тела, вызывают уменьшение массы головного мозга, а также установлена положительная корреляционная связь массы тела и мозга у новорожденных, которая проявляется как у человека, так и у экспериментальных животных (7).
Причинами низкой массы тела при рождении указываются качественная и количественная неполноценность питания матери во время беременности, многоплодная беременность, различные болезни матери, нарушающие развитие плаценты, снижающие ее транспортные функции. Во всех описанных ситуациях плод испытывает дефицит питательных веществ, который может снижать интенсивность синтетических процессов в нейронах, скорость роста их отростков. Кроме того, к снижению массы тела плода могут вести также воздействия радиации, алкоголя, наркотиков, лекарственных препаратов, вирусов и бактерий. Эти агенты могут приводить к поражению плаценты, уменьшать ее способность доставки необходимых компонентов от матери к плоду и удаления шлаковых продуктов последнего, то есть эффект их воздействий также может реализовываться через уменьшение количества поступающих питательных веществ. Перечисленные факторы обладают способностью и прямо повреждать клетки развивающегося мозга. Исследованиями в области гормонального фона установлено, что гормоны, циркулирующие в крови плода, синтезируются плацентой, эндокринными железами матери и его собственными железами. Рост, развитие и функционирование желез внутренней секреции уже в эмбриогенезе в значительной степени регулируются гипоталамусом (7).
Также установлено, что гормональные воздействия оказывают:
- необратимое, программирующее, изменяющее фенотип и функциональную активность клеток влияние,
- обратимое, регуляторное, изменяющее клеточные функции (7).
Влияние концентрации гормонов в крови матери на мозг плода реализуется различными путями:
- часть гормонов (стероидные), продуцируемые корой надпочечников, яичниками, проходит через плаценту в кровь плода, где они прямо воздействуют на нейробласты, нейроны и другие клетки мозга;
- гормоны гипофиза, яичников, надпочечников и других желез матери влияют на развитие плаценты, определяя эффективность транспорта различных компонентов от матери к плоду и в обратном направлении;
- изменения эндокринных функций плаценты, вырабатывающей большое число различных гормонов, поступающих как к матери, так и к плоду, также могут оказать воздействие на формирование мозга;
- изменения содержания гормонов в крови матери и плода влияет на развитие его мозга и опосредованно, воздействуя на концентрацию в крови таких важных для этого процесса веществ, как глюкоза, аминокислоты, разные классы липидов и т.д.;
- при снижении функции желез у матери железы плода, стремясь компенсировать дефицит соответствующих гормонов, находятся в состоянии гиперфункции, а следствием этих процессов является то, что после рождения характер развития соответствующих желез, их функциональная активность, реакция на стимулирующие воздействия отличаются от контрольных, что немаловажно, в части показателей высшей нервной деятельности у животных с измененными вследствие указанных выше причин эндокринными железами (7).
Приняв к сведению вышеизложенное, рассмотрим некоторые фрагменты существующих ныне технологий, сопряженных с процессами воспроизводства быстроаллюрных лошадей.
“Случку при конюшенном содержании проводят с 15 февраля по 15 июля, при табунном – с 15 апреля по 1 августа. Эти сроки могут несколько изменяться в зависимости от климатических условий. Во всех хозяйствах целесообразно проводить случку кобыл, по возможности, в более ранние сроки, чтобы окончить выжеребку до начала весенних полевых работ и чтобы жеребята к отъему нормально развились” (5-стр.99).
По нашему мнению, зависимость технологии современного быстроаллюрного коневодства от климатических и иных сезонных факторов представляет серьезное препятствие по организации интенсивного и расширенного воспроизводства конского поголовья из-за частых прохолостов кобыл, возникающих по различным причинам и как следствие этого – понижение зажеребляемости и выхода жеребят. Сезонность выжеребки в некоторой степени также оказывает влияние на сезонную потребность, определенными технологиями конезаводства, в квалифицированных кадрах. По этим причинам, снятие воздействия сезонных факторов на современные технологии коневодства - одна из проблем, требующих своего инженерно-технического и организационно-правового решения.
Инженерно-технические решения, по нашему мнению, должны акцентироваться на вопросах экологии содержания, тренинга и кормления лошадей. В то время как организационно-правовые решения должны направляться на снятие каких-либо регламентирующих, как и других фактически существующих ограничений, обусловленных сроками выжеребки быстроаллюрных лошадей.
Одним из заблуждений, доставшихся нам из прошлого, представляется нам недооценка значимости маточного поголовья в сравнении с жеребцами-производителями, что неминуемо отражается на качестве формируемого маточного состава. В доказательство данного опасения приводим некоторые выдержки из руководств.
“Матки из-за малоплодности не могут оказать такого влияния на структуру пород, как производители...” (2).
“Согласно наблюдениям, жеребчики более требовательны к условиям кормления и содержания, чем кобылки, и значительно лучше отзываются на полноценное кормление и хорошее содержание.… Поэтому нормы кормления жеребчиков должны быть несколько выше норм кормления кобылок” (4-стр.122).
По нашему мнению, различия в нормах кормления жеребчиков и кобылок не означают большую или меньшую требовательность к качеству рационов, а свидетельствуют о количественной разнице в потребностях, обусловленной физиологическими различиями в росте и развитии, и никаким образом не означает различия в прихотливости. А вместе с тем степень развития будущих племенных кобыл, куда сильнее и глубже может повлиять на качество их будущего приплода, множеством факторов (см. выше), нежели степень развития будущих жеребцов-производителей.
Жеребость – сложный физиологический процесс, требующий высококвалифицированного подхода для обеспечения правильных условий кормления, содержания и проведения моциона кобыл.
Таким образом, влияние условий содержания племенных лошадей имеет качественную и количественную составляющие.
Качественная составляющая заключается в улучшении состояния здоровья получаемого приплода, а также улучшении показателей развития и роста жеребят и как следствие, возможности выращивания более ценного поголовья быстроаллюрных лошадей.
Проблематика коневодства, связанная с периодом развития и роста жеребят, требует от зоотехнических специалистов комплексного подхода, выражаемого в научно обоснованных и ясных решениях. Под ясностью решений нами подразумевается, прежде всего, однозначность и понятность способов достижения цели, поставленной перед технологией её преследующей.
Далее приведем выдержку о том, как предлагается понимать решение задачи кормления жеребых кобыл применительно к существующей практике.
“Забота о жеребенке должна начинаться, когда он еще в утробе матери. Жеребых кобыл обеспечивают полноценными кормами: сеном, концентратами, корнеплодами, кормами, богатыми витаминами (особенно каротином) и минеральными веществами. В первой половине жеребости, когда формируются ткани и органы зародыша, неполноценное питание кобыл, прежде всего недостаток в кормах витаминов и микроэлементов, часто приводит к скрытым абортам. Во второй половине жеребости в развивающемся плоде интенсивно нарастает масса скелета; у новорожденного жеребенка она составляет 31% его живой массы. Резко увеличивается потребность жеребой кобылы в кальции и фосфоре. В последние месяцы жеребости усиливается нарастание мышц плода и увеличивается потребность в белковых веществах, поэтому так важно полноценное протеиновое кормление” (5- стр.104).
По нашему мнению, термин “полноценный корм” означает покрытие потребностей организма необходимым количеством переваримых веществ, содержащихся в корме. Однако, применение полноценного корма не равнозначно с понятием комфортного кормления, прежде всего ввиду того, что комфортное кормление предполагает ещё, сбалансированное кормление и даже не только это. При этом надо ещё учитывать степень эксплуатации (интенсивность работы) лошади. В свою очередь, степень эксплуатации определяется величиной селекционируемого человеком хозяйстсвенно-полезного признака в заданный момент времени и в конкретных условиях. Таким образом, рацион племенной быстроаллюрной лошади должен быть обеспечен, как переваримыми веществами в свободном и связанном виде, так и непериваримыми (балластными), в количествах минимально возможных, достаточных и необходимых для обеспечения всех трофических потребностей организма лошади от одного приема корма до другого с учетом индивидуальных особенностей животного, в том числе времени необходимого до начала поступления в кровь лошади новой порции питательных веществ от новой порции корма и т.д.
По нашему мнению, принцип сбалансированности и адекватности, соответствующему режиму продуктивности животного, корма для удобства реализации в конкретных рационах, необходимо дифференцировать по группам питательных веществ, разделенных по степени значимости для обеспечения трофических потребностей:
- основные питательные вещества (переваримые протеин, жиры, углеводы);
- протеин (соотношение аминокислот
);
- углеводы (соотношение БЭВ, легко- и труднопереваримой клетчатки);
- жиры и масла (соотношение жирных кислот);
- минеральные вещества (соотношение макро- и микроэлементов);
- биологически активные вещества (соотношение витаминов).
Уровень организации кормления во многом определяется и квалификацией обслуживающего лошадей персонала.
Учитывая узкий набор кормовых средств, традиционно применяемых для кормления лошадей быстроаллюрных пород (овес, сено, шрот, меласса, сахар, травяная мука) и невозможность замены сбалансированного рациона кормовыми добавками, которые не решают проблемы лишних (а часто и вредных из-за переизбытка) веществ и понижают общую переваримость рациона, можно утверждать, что кормлению лошадей повсеместно уделяется неоправданно мало внимания.
Глава 2. Технологии выращивания и содержания быстроаллюрных лошадей.
2.1. Влияние условий среды и физической нагрузки на организм животных
Способность живых существ приспосабливаться к меняющимся условиям среды – одна из их важнейших особенностей, позволяющих организмам существовать в самых различных условиях внешней среды.
Здоровье лошади зависит, главным образом от условий во внешней и внутренней средах, факторы и свойства которых, воздействуя на организм лошади в качестве раздражителей, определяют естественную реакцию организма лошади на эти воздействия, и последующую адаптацию к данным условиям.
Лошади, как и многие другие животные, дышащие атмосферным воздухом, находятся в постоянном взаимодействии с воздушной средой. По мере эволюции вида, природа выработала у лошадей ряд качеств, отвечающих на воздействие факторов окружающей среды. Так температуре окружающей среды противостоит терморегуляция, а влажности воздуха – транспирация и перспирация.
К наиболее важным факторам окружающей среды принято относить следующие:
1. Температура воздуха;
2. Газовый состав воздуха;
3. Влажность и скорость воздуха;
4. Давление воздуха;
5. Аэроионизация;
6. Пылевая и микробная загрязненность воздуха;
7. Солнечная радиация;
8. Электрические и электромагнитные поля;
9. Шум.
В процессе эволюции видов в каждой климатической зоне появляются виды животных соответствующих именно данному набору параметров среды их обитания.
Так в жарком климате у лошадей тонкая кожа с редкими, короткими и тонкими волосами, слаборазвитая подкожная клетчатка, сухой тип конституции, слаборазвитые челка, хвост и щетки. В тоже время лошади северной зоны имеют ярко выраженную способность к быстрому жироотложению (2).
Температурная адаптация лошадей сопровождается совокупностью специфических физиологических изменений, главными из которых, по мере пребывания в условиях повышенной температуры среды, являются усиление потоотделения, снижение температуры ядра и поверхности тела, уменьшение частоты сердечных сокращений при нагрузке и увеличение числа секретируемых потовых желез. Механизмы температурной адаптации направлены на усиление отдачи тепла организмом лошади во внешнюю среду. При любом увеличении температуры организма лошади, в частности и во время работы, потоотделение начинается быстрее, т.е. снижается температурный порог потоотделения и увеличивается его скорость (35).
После адаптации организма к жаре меньше пота стекает в виде капелек, не испаряясь, так как пот более равномерно распределяется по всей поверхности тела, чем у неадаптированного животного, т.е. возрастает площадь поверхности кожи, охваченной для усиленной теплоотдачи испарением. Как следствие, усиление потоиспарения ведет к более эффективному снижению температуры кожи, благодаря чему кровь кожных покровов охлаждается быстрее, усиливается физический транспорт тепла от глубоких частей тела и более нагретых к его поверхности (35).
Главным эффектом усиления адаптивных температурных механизмов является снижение температуры тела, как в условиях покоя, так и её прирост в момент мышечной работы (35), увеличивается температурный диапазон мышечной работы, а также снижается тепловая нагрузка в легких из-за снижения суммарного тепла реакций оксигенации, что в свою очередь ведет к увеличению коэффициента полезного действия (к.п.д.) мышечных волокон.
В течение температурной адаптации снижается содержание солей в поте, увеличивается выделение воды и как следствие, концентрация электролитов в крови повышается, что рефлекторно вызывает сильное ощущение жажды, являющееся механизмом, направленным на компенсацию потерь воды организмом животного. У неадаптированного к жаре организма данный рефлекс хуже выражен, что может приводить к обезвоживанию организма, а при резком водопотреблении и к опою. Адаптированное к жаре животное имеет четко выраженный рефлекс для поддержания водного баланса организма (35).
По мере адаптации к высокой температуре снижается проницаемость капилляров кожного покрова, уменьшается выход молекул белка из капилляров, содержание белка в тканевой жидкости кожи увеличивается. При тепловых воздействиях белок интенсивно перемещается транзитно через лимфатическую сеть кожи в кровь, что позволяет сохранять её высокие онкотическое давление и объем. Объем циркулирующей крови в состоянии покоя увеличивается, а показатель гематокрита и вязкость крови имеют тенденцию к снижению (35).
Температурная адаптация приводит к снижению нагрузки на сердечно-сосудистую систему, к росту возможности эффективного усиления кожного кровотока, за счет более быстрого перемещения крови в систему кожных капилляров, а за счет раскрытия и расширения сети поверхностных капилляров, к приближению крови к поверхности тела животного и более эффективному её распределению (35).
По мере протекания процесса температурной адаптации уменьшается степень сужения сосудов в чревной и почечной областях, что улучшает кровоснабжение органов брюшной полости во время физической работы, повышается механическая эффективность выполнения физической работы в условиях повышенной температуры, сопровождаемая прогрессивным снижением потребления кислорода при выполнении работы, снижается частота сердечных
сокращений в покое и при работе, увеличивается систолический объем, что не изменяет сердечный выброс на протяжении периода пребывания в жарких условиях, а также увеличивается венозный возврат, за счет повышения объема циркулирующей крови (35).
Тренировочные и состязательные нагрузки, требующие проявления выносливости, вызывают существенное повышение температуры ядра тела – до 40
оС, даже в нейтральных условиях среды, что служит стимулом для развития адаптационных реакций к повышенной тепловой нагрузке с морфологическим оформлением ответа на эти реакции. Данные реакции, со стороны сердечно-сосудистой системы, потовых желез и других органов, во многом сходны с реакцией организма животного на акклиматизацию к жарким условиям (35).
В результате систематических физических нагрузок у тренируемых животных совершенствуется терморегуляция:
- снижается теплопродукция;
- повышается чувствительность реакции потоотделения на тепловое раздражение, выражающееся в сокращении времени от начала теплового воздействия до начала потоотделения;
- увеличивается объем циркулирующей крови;
- совершенствуются реакции перераспределения кровотока за счет раскрытия-сужения сосудов и подключения в работу закрытых сосудов кожного покрова;
- более равномерное распределение потоотделения по поверхности тела, за счет увеличения числа функционирующих потовых желез;
- понижается концентрация солей в поте;
- улучшается способность к теплопотерям за счет повышенного потообразования;
- понижается внутренняя и кожная температура тела (35).
Вышеизложенное морфологическое оформление ответа на реакции к повышению температуры и к физической нагрузке особенно важно учитывать с момента рождения жеребенка и последующей адаптации к условиям внешней среды, при этом важным моментом совершенствования его организма в связи с последующей специализацией использования, является как возможно ранняя организация тренировочного процесса с целью грамотного задания адекватной степени развития организма жеребенка физической нагрузки.
Оптимальному значению каждой из переменных внутренней среды организма (температура тела, рО
2 в тканях и т.п.) соответствуют диапазоны значений внешних факторов природы. В случае отклонения от этих значений включается физиологическая регуляция, направленное на поддержание гомеостаза, на расширение и смещение диапазона устойчивости организма. Главенствующее положение в системе адаптации организма занимает установление оптимизированного стабилизированного уровня затрат энергии и в связи с этим, снижение энергетической цены гомеостаза становится основным принципом физиологической адаптации к воздействию внешних факторов.
В качестве естественных разобщителей окисления и фосфорилирования выступают тиреоидные гормоны, повышающие теплопродукцию мышечного сокращения (24-стр.128).
При хроническом действии низкой температуры в мышцах происходит увеличение относительного количества медленных (красных) и промежуточных волокон, для которых характерен изометрический тип мышечных сокращений, а также происходит повышение потребления медленными (красными) волокнами кислорода, что усиливает гипоксию при двигательной деятельности (15-стр.116). Кроме того, при длительном воздействии низких температур происходит угнетение функций физической терморегуляции, что одновременно понижает эффективность мышечной работы (к.п.д. мышц), которая напрямую зависит от эффективности системы теплоотдачи (системы “охлаждения” организма).
К.П.Д. мышц – представляет собой отношение кинетической энергии мышечного сокращения ко всему количеству энергии, вырабатываемой мышцами по время своей работы. Постепенно, к.п.д. тренированной мышцы повышается и может достигать 50% (18-стр.17).
В условиях покоя мышцы потребляют 20-25% всей энергии, которую производит организм. В обычных условиях мышечная масса вырабатывает 50-60% тепла организма. При усиленной работе этот показатель достигает 80%, поэтому эволюцией видов мышцам отведена ещё одна важнейшая функция – участие в терморегуляции (18-стр.11). Во время работы одна часть энергии превращается в тепло, а другая – преобразуется в механическую работу, при этом соотношения между этими частями отражает к.п.д. мышечных сокращений. При этом, учитывая тот факт, что сократительный белок не может выполнять свою функцию в условиях денатурации, к.п.д. мышц отчасти отражает уровень готовности мышечной ткани к выполнению физическую нагрузки максимальной мощности.
В процессе эволюции гомойотермные животные выработали способность использовать двигательную активность для защиты от холода, как во взрослом состоянии, так и в новорожденном, когда эта способность наиболее ярко выражена. На основании литературных данных и по научным исследованиям сделано утверждение, что “… все случаи химической терморегуляции сочетаются с повышением сократительной активности мышц. Это – двигательная активность, холодовая дрожь, терморегуляционный тонус. Мощность их довольно велика” (17-стр.39).
Также было установлено, что при акклиматизации к холоду химическая терморегуляция осуществляется при низком уровне электрической активности мышц. Это навело ученых на мысль, что “… после акклиматизации к холоду теплопродукция мышечных сокращений существенно повышается” (17-стр.40) и этот факт был подтвержден прямым определением потребления кислорода мышцами бедра акклиматизированных и контрольных кроликов. Исследования, проведенные на изолированной диафрагме крыс, показали увеличение теплопродукции, рассчитанной на 1 г массы и 1 г развиваемой силы, у акклиматизированных к холоду животных на 38% и 54% соответственно больше, чем у контрольных (17-стр.41). Что касается механизма уменьшения к.п.д. мышц, то
учеными-исследователями делается вывод, “…что множество даже очень слабых сокращений, которые не производят практически никакой механической работы, могут стать источником значительного количества тепла мышечной ткани. Но именно такая деятельность имеет место в холодовой мышечной дрожи или при так называемом терморегуляционном мышечном тонусе, состоящих из частых, слабых, нерегулируемых сокращений отдельных групп мышечных волокон” (17-стр.45), а механизмом понижения к.п.д. мышечной деятельности и повышения теплопродукции может являться разобщение окисления и фосфорилирования (17-стр.46).
Относительно уровня понижения к.п.д. мышц, исследованиями на мышцах задних конечностей кролика установлено, что после акклиматизации к холоду к.п.д. мышц падает в
1,5 – 2 раза, при этом эффективность терморегуляционного тонуса повышается на 200-300%, а прирост теплопродукции организма в целом может составить 90-135% (17-стр.48). На основании полученных данных делается вывод о том, что понижение к.п.д. сократительной деятельности мышц является важным механизмом повышения уровня теплопродукции организма. Подчеркивается также тот факт, что речь идет о понижении к.п.д. мышц не на доли процентов, а в 1,5-3 раза!!! (17-стр.49), аналогичные данные подтверждаются и в других исследованиях (19-стр.13).
Также установлено, что адаптированные к холоду животные по сравнению с адаптированными к теплу имеют более высокий уровень метаболизма, как в тепле, так и в холоде, что выделяет процесс химической терморегуляции в категорию конкурирующего процесса по потреблению кислорода по отношению к мышечной деятельности. Кроме того, проведенные опыты на амбарных крысах определенно указывают на существенное изменение энергетики мышечного сокращения при адаптации животных к холоду, приводящее к увеличению тепловыделения при элементарном сократительном акте (15 – стр.113), что безусловным образом, сказывается на понижении эффективности мышечного сокращения (к.п.д. мышц). При метаболической адаптации к холоду изменяется энергообразование в мышечных волокнах. При этом повышается эффективность отставленного теплообразования за счет снижения к.п.д. химической работы по ресинтезу АТФ (24-стр.128).
При адаптации к холоду происходит интенсификация обмена липидов, возрастает их концентрация в крови и в клетках многих тканей. Липидная перестройка организма вызывается повышением потребности в липидах с одновременным изменением качественного состава липидов. Изменяется соотношение между триглицеридами, фосфолипидами, стеринами, увеличивается доля и концентрация высокомолекулярных (трудноизвлекаемых) жирных кислот, выполняющих роль главного субстрата окисления (24-стр.131), таким образом, повышается плотность жировой ткани и понижается её эластичность и она начинает оказывать ещё большее сопротивление мышечному сокращению. Мышечные волокна не имеют собственных адренорецепторов (преимущественно
a-адренорецепторы), симпатические окончания в мышцах локализованы преимущественно в стенках капилляров и перикапиллярных соединительнотканных образованиях (мышечный жир), расположенных внутри скелетных мышц, а норадреналин индуцирует образование и доставку к мышечным митохондриям свободных жирных кислот.
По современным представлениям, в мышцах млекопитающих выделяют 2 основных типа волокон.
Красные (медленные) – малого диаметра, для обеспечения более эффективного обмена веществ и тепла, они темные, хорошо снабжаются кровью, в них много миоглобина и мало гликогена, к тому же обладают низкой способностью к его расщеплению, содержат большое количество митохондрий, нейтральных жиров и ферментов для обеспечения липидного и окислительного обмена.
Белые (быстрые) – светлые, большого диаметра, содержат мало миоглобина, бедно снабжаются кровью, богаты гликогеном, содержат большое количество гликолитических ферментов и обладают высокой способностью гликогенолиза и гликолиза (превосходящая красные волокна), у них слабая способность к липидному и окислительному обмену. Гликоген мышц служит резервом синтеза АТФ внутри мышечной ткани, для необходимости многократного повышения мышечной активности (
38-стр.158) в результате анаэробного ресинтеза АТФ. При выполнении мышечной работы максимальной напряженности анаэробный путь ресинтеза АТФ обеспечивает на 80-90% общую потребность мышц в АТФ, поскольку гипоксия резко возрастает и сердечно-сосудистая система только ещё начинает увеличивать доставку кислорода к работающим мышцам для удовлетворения их метаболических потребностей (37-стр.10).
Выделяют ещё 2 типа волокон с промежуточными свойствами.
Одни волокна с оксидативно-гликолитическим обменом, а другие обладают обменом, приближающимся к гликолитическому, хотя частично имеют и оксидативный обмен.
Высокоинтенсивные физические нагрузки в пределах 3-х минут повышают активность миозина, Са
2+ и Na+-K+-АТФзы, которые осуществляют гидролиз АТФ с высокой скоростью (37-стр.10).
АТФазы
АТФ <--> АДФ+ Ф
н+Н+
где, АДФ – аденозин 5-дифосфат, Ф
н– неорганический фосфат.
Последовательность, в которой происходит анаэробный ресинтез АТФ, при возрастающей потребности в нем, выглядит таким образом (37-стр.10) (Рис.2):
КФК
КФ+АДФ+Н
+ <--> АТФ+Кр
Гликоген+3АДФ+3Ф
н <--> 3АТФ+2лактат-+2Н+
АК
2АДФ <--> АТФ+АМФ
АМФдезаминаза
АМФ+ Н
+ <--> ИМФ+NH4+,
где: КФ – креатинфосфат, К
р - креатин, КФК – креатинфосфокиназа, АК – аденилаткиназа, АМФ – аденозин 5-монофосфат, ИМФ – инозинмонофосфат.
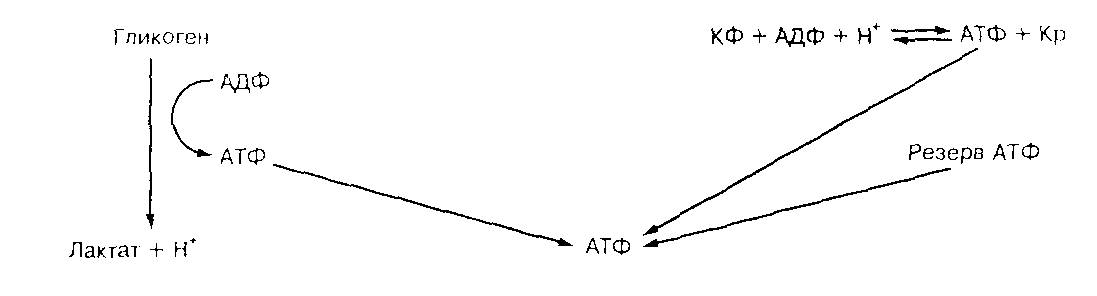
Рис.2 Схема источников анаэробного ресинтеза АТФ.
Распад КФ происходит очень быстро, т.к. он используется как субстрат для ресинтеза АТФ на самой начальной стадии интенсивной физической нагрузки (до 10 с). В то же время с началом интенсивной физической нагрузки активизируется и анаэробные гликолиз и гликогенолиз (37-стр.12-13). Скорость обеспечения ресинтеза АТФ анаэробными путями, является предельной скоростью для осуществления мышечной деятельности высокой мощности и может достигать 2-4 раза большую, чем соответствующую максимальному уровню потребления кислорода (МПК). При выполнении максимальной мышечной деятельности более 10 с, мощность выполняемой работы снижается, а скорость гликолитического и КФ-зависимого ресинтеза АТФ не будет максимальной (37-стр.13). Самые высокие скорости ресинтеза АТФ за счет КФ и гликолитических процессов при физических нагрузках высокой интенсивности проявляются в первые 10 с (Рис.3). При продолжении таких физических нагрузок до 30 с, запасы КФ
истощаются, а скорость гликолитического ресинтеза АТФ будет составлять около 50% скорости ресинтеза в первые 10 с.
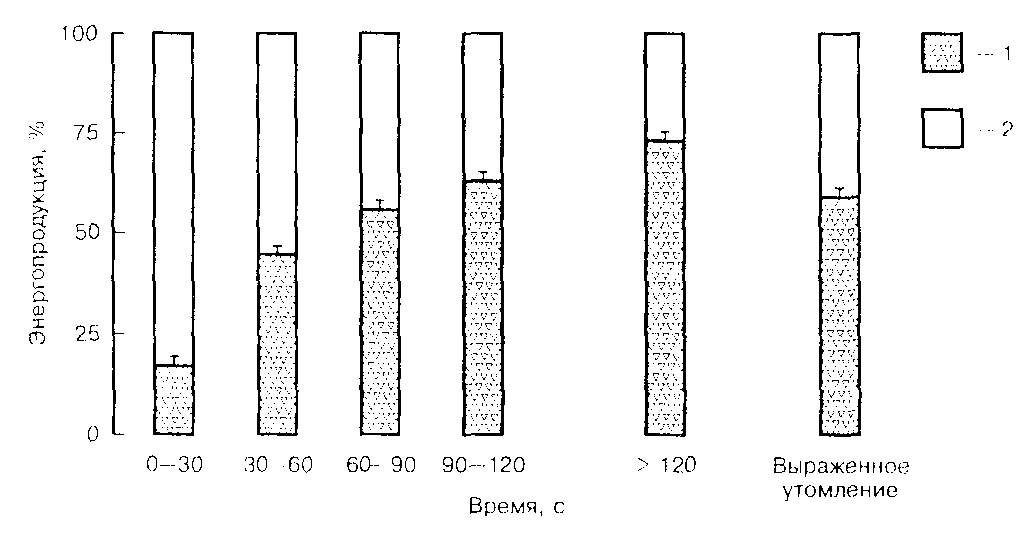
Рис.3 Относительный вклад аэробного (1) и анаэробного (2)
ресинтеза АТФ на протяжении 30-секундных интервалов
предельной (около 3 мин) физической нагрузки
При этом гликолитическое обеспечение АТФ при 30-секундной максимальной физической нагрузке выше в 3…4 раза, чем при обеспечении только за счет КФ-механизма (37-стр.17). Это говорит о том, что тренировочный эффект (за счет развития процессов гликолитического и КФ-механизма ресинтеза АТФ) при тренинге быстроаллюрной лошади будет максимальным, если интервалы максимальных физических нагрузок не будут продолжительностью более 30 с, во избежание развития конкурентоспособности энергообразования аэробного процесса перед анаэробным.
Так в результате тренировки бегунов средний показатель мощности работы при 30-секундном беге с максимальной скоростью увеличился на 60%, а анаэробная продукция АТФ за счет гликолиза выросла на 20%, тогда как вклад энергопродукции за счет КФ и запасов АТФ практически не изменился. Кроме того, имеются литературные данные о том, что под влиянием 4…6 недельной тренировки в велосипедном спринте снизилось содержание красных (медленных) волокон с 57% до 48% и увеличилось содержание белых (быстрых) волокон с 32% до 38% (37-стр.31-32).
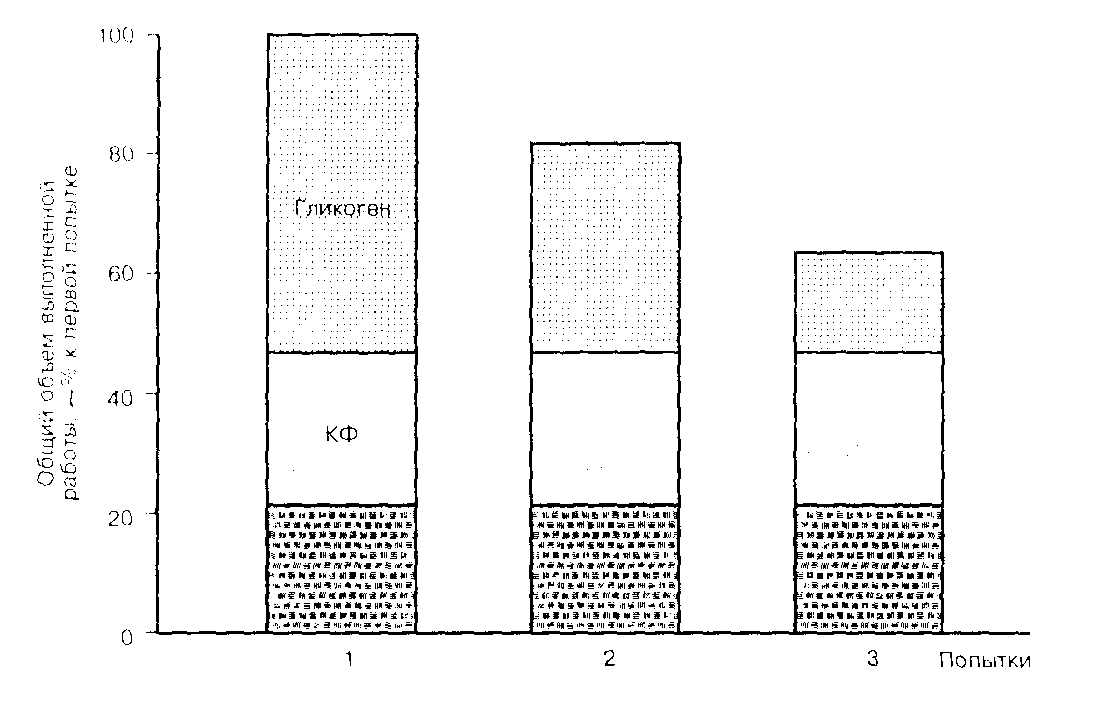
Рис.4 Участие отдельных энергетических механизмов в общей
энергопродукции при трёх максимальных нагрузках продол-
жительностью 30 с каждая с 4-минутными периодами отдыха.
При грамотном применении тренировочных нагрузок, повышение работоспособности скелетных мышц достигается увеличением содержания в них КФ до начала физической нагрузки, а также за счет повышенной способности к его ресинтезу в периоды отдыха. Анаэробные же гликолитические возможности повышаются за счет восстановления истощенных нагрузкой высокой интенсивности запасов в период пониженной мышечной деятельности на фоне высокоуглеводного питания (37-стр.36-37) (Рис.4).
Имеются литературные данные о том, что при адаптации к холоду происходит уменьшение доли белых (быстрых) волокон и увеличение доли красных (медленных) и промежуточных волокон мышечной ткани, а состояние Т-системы (продолжительность следового отрицательного потенциал сопровождающего спайк, пик потенциала действия, ионная проницаемость мембран) белых и красных мышечных волокон стремится к промежуточному состоянию, что выражается проявлением процессов нейрогуморальной координации и дифференциации терморегуляционной активности мышц. Именно для медленных волокон характерны более высокие уровни аэробных процессов, активности окислительных ферментов, концентрации многих энергетических субстратов и миоглобина, лабильность системы окислительного фосфорилирования и её реактивности по отношению к разобщителям (15-стр.85). Так холодовая адаптация белых крыс (4-5 недель)сопровождается заметным повышением активности сукцинатдегидрогеназы (СДГ) в красных волокнах мышцы бедра и особенно в четырехглавой (на 40%). Увеличение активности СДГ происходит почти целиком за счет красных волокон “ядра” мышцы, в которых увеличивается и число митохондрий. “Красное ядро” четырехглавой мышцы контрольных животных составляет 35-38%, а у адаптированных – 47-58% её общего веса, причем разница возрастает с увеличением срока адаптации. Увеличение активности СДГ связывают с повышением мощности сукцинатоксидазной системы, что продиктовано потребностью повышенного окисления сукцината, который в отличие от НАД-зависимых субстратов быстро и многократно увеличивает скорость обмена (20-стр.14-17). Вместе с тем, адаптивные изменения к холоду в мышечных волокнах разных типов остаются малоизученными.
Закономерности различий размеров тела животных одного и того же вида в связи с различиями климатических условий сводятся к тому, что в холодных условиях животные более крупные, отношение единицы поверхности тела к массе тела меньше, размеры выступающих частей тела уменьшены, кроме того, обеспечены более толстыми и эффективными теплоизолирующими средствами (кожный покров, волос, подкожная жировая ткань), чем животные того же вида в теплых климатических зонах (16 – стр.12). Изменения кожного и волосяного покровов организма лошади являются проявлением адаптации вида к условиям внешней среды зоны обитания. Основную функцию в теплозащите от холода путем снижения конвекционных потерь тепла выполняет воздух, находящийся между волосами и в их сердцевинном слое. Стремление к препятствованию потерь тепла заставляет организм лошадей увеличивать число волос, удлинять их длину, развивать сердцевинный слой волос, увеличивать толщину кожного покрова и отложения подкожного жирового слоя (16).
Подключение функциональных резервов для целей адаптации к низким температурам меняет внутреннюю среду организма и включает механизм перестройки для целей выживания (увеличение эритроцитарной массы, объема крови, её компонентов и энергетических депо)(16), понижая полезный потенциал организма на выполнение максимальной физической нагрузки.
Атмосферный воздух содержит промышленные выбросы, но наиболее опасным для организма лошади может стать воздух в конюшне.
Наиболее опасными для лошадей нам представляются соединения азота. Основным источником их появления в воздухе являются разлагающиеся кал, моча и подстилка. В зоотехнической литературе до сих пор присутствует, наряду с другими способами хранения навоза, способ хранения навоза под скотом. Убогость и порочность данной методики кроме как недоумения, ничего более достойного не вызывает.
Все оксиды и закиси азота в соединении с гемоглобином крови составляют метгемоглобин, что приводит к нарушению доставки кислорода к органам и тканям, выделения углекислоты из организма и приводит к приступам асфиксии. Отмечено действие таких соединений на центральную нервную систему. Действие данных соединений безусловным образом тормозит метаболические процессы в организме и вместе с этим, снижает качество полезных признаков животных (резвость, качество и длину прыжка и т.п.).
Аммиак образуется при разложении органических азотсодержащих соединений под действием уреазоактивных анаэробных бактерий. Наиболее высокая концентрация его наблюдается вблизи пола. Аммиак – агрессивный газ. Вызывает сильное раздражение верхних дыхательных путей, конъюнктивы и роговицы глаз, изо рта и носа выделения пенистой жидкости, иногда с примесью крови, нарушение свертываемости крови в результате прямого действия на протромбин, повышение содержания остаточного N
2 в крови за счет быстрого накопления в ней NH3, а также вызывает лейкоцитоз, уменьшение содержания в крови и печени общего и свободного витамина В1. Аммиак рефлекторно вызывает остановки дыхания и кровообращения, могущие привести к смерти. Окись углерода (СО2) усиливает действие аммиака (10). При его вдыхании возможен ожог слизистой оболочки. Уже в малых дозах он парализует деятельность ворсинок дыхательных путей, как щелочь, он подщелачивает кожу и копытный рог, разрыхляя их. При попадании на незащищенный влажный кожный покров, особенно конечности, он под действием кислорода превращается в нитрит – чрезвычайно ядовитое соединение, вызывая цианоз кожных покровов, отставания в росте, отечность и кровоизлияния в миокарде, селезенке, печени, легких и мозге. При наличии аммиака в воздухе у лошадей наблюдаются конъюнктивиты, слезотечение и т.п. Аммиак в крови животных при соединении с гемоглобином образует щелочной гематин и даже метгемоглобин (через нитриты), вызывая анемии и приводит к снижению полезных качеств животных. Кроме того, аммиак очень хорошо адсорбируется стенами и другими влажными и пористыми поверхностями. Адсорбция пропорциональна концентрации аммиака и возрастает с повышением влажности и снижением температуры. При увеличении температуры аммиак выделяется в воздух, что обостряет вышеприведенные действия на организм лошадей и усугубляет эпизоотическую обстановку по заболеваниям слизистых оболочек, кожи и дыхательных путей (гнойные, иногда геморрагическое воспаления горла, бронхов и легких; фибринозный или гнойный плеврит). При очень больших концентрациях (1-3 мг/л) аммиака может наступить смерть от отека легких (9).
Из курса физиологии нам известно, что выделение аммиака из организма животных происходит не только через мочу. Но и через легкие, благодаря разности напряжения аммиака в крови и в альвеолярном воздухе. Чем выше эта разность. Тем интенсивнее скорость выведения аммиака из организма животного и как следствие понижаются затраты АТФ организмом на связывание аммиака в карбамид. Таким образом, снижение содержания в воздухе конюшни аммиака, через уменьшение расхода АТФ, повышает общую работоспособность лошадей.
Дополнительно следует отметить, что под воздействием постоянно действующей концентрации аммиака, возникает адаптационная реакция организма лошади к данной концентрации путем стабилизации метаболических процессов (в основе лежит способность организма к образованию новых временных рефлекторных связей, способствующих выработке наиболее целесообразной адаптационной ответной реакции), что приводит к невозможности резкого изменения в определенный момент (особенно во время скачек и соревнований – когда это наиболее нам необходимо) в сторону ускорения метаболических процессов организмом лошади “ни кнутом, ни пряником”, именно потому, что данные процессы основаны на временных рефлекторных связях.
Предельно допустимой концентрацией аммиака в конюшнях почему-то считается 5-20 мг/м
3 в зависимости от возраста лошади. Однако в данной дипломной работе мы обсуждаем традиционные технологии быстроаллюрного коневодства, т.е. имеем в виду технологии содержания самых дорогих животных на планете. По этой причине технологии содержания этих животных должны отвечать требованиям и быть наиболее эффективными для удовлетворения ожидаемого нами качества быстроаллюрных лошадей. Исходя из вышеизложенного, следует заключить, что любое заметное нахождение аммиака в воздухе конюшни, мы должны рассматривать как чрезвычайную ситуацию.
Высокое содержание углекислого газа также увеличивает накопление продуктов метаболических реакций, в том числе и недоокисленных, в крови лошади, главным образом, из-за выравнивания парциальных давлений углекислого газа в альвеолярном воздухе и в крови.
Одним из наиболее опасных газов, сопутствующих традиционным технологиям конезаводства, является сероводород (H
2S), возникающий при гниении белковых серосодержащих веществ и в кишечных газах. Сероводород:
- блокирует ферментативные процессы;
- за счет своей кислотности, раздражает слизистые оболочки, геморрагические воспаления верхних дыхательных путей и легких, отек легких, изъявления роговицы;
- воздействует на центральную нервную систему, вызывая нарушения условнорефлекторной деятельности;
- в результате окисления его в тканях, вызывает образование перекисных соединений, угнетающих гликолиз;
- вызывает общее отравление организма.
Из вышеизложенного следует заключить, что основным фактором, влияющим на газовый состав конюшенного воздуха, является подстилка. По этой причине, помимо традиционных мероприятий по обеспечению снижения и стабилизации содержания вредных газов:
- своевременного удаления кала лошадей и мокрой подстилки, применения влагонепроницаемых полов, эффективного вентилирования конюшен, использования газопоглощающих материалов, обеспечения работы канализации,
- все большее внимание следует уделять разработке новых технологий содержания лошадей, где образование потенциально вредных факторов, в частности из подстилки, будет сведено к минимуму. Так эффективным решением данной проблемы может стать применение дернины в денниках в качестве несменяемой подстилки. Эффективность внедрения данного технического решения в практике зависит от ряда параметров и наличия систем:
- размер денника – 60-80 м
2 (влияет на пастьбоустойчивость дернины),
- грамотный подбор растений для устройства дернины, имеющих различную пастьбоустойчивость,
- температурно-влажностного режим в деннике 18-26
ОС (активный рост большинства трав начинается при температуре выше 15ОС) зависит от возраста, породы лошади и степени её адаптации к повышенной температуре,
- совмещение или разделение мест мочеиспускания и скаливания,
- освещенность и облучение УФ-облучателями денника,
- устройство дренажа и системы пролива водой мест мочеиспускания и др.
Учитывая вышеуказанные факторы и параметры можно предложить два варианта системы комфортного содержания лошадей:
- изолированные денники со смежными уборными помещениями для мочеиспускания и дефекации лошадей (Рис.5),
- изолированные денники со смежными зонами для мочеиспускания и дефекации лошадей (Рис.6).

Рис.5 Схема фрагмента конюшни с изолированными денниками (1) со смежными уборными помещениями (2) для мочеиспускания и дефекации лошадей
.
3 – помещение для амуниции,
4 – проход.

Рис.6 Схема фрагмента конюшни с изолированными денниками (1) со смежными зонами (2) для мочеиспускания и дефекации лошадей.
4 – проход.
Принимая во внимание тот факт, что лошади, как и многие другие животные склонны к мечению своей территории, можно предложить устраивать места для скаливания и мочеиспускания смежно и попарно у денников с лошадьми одного пола, разделяя данные зоны (или помещения) решетчатыми заграждениями до пола помещения для того, чтобы лошади могли видеть по весь рост и чувствовать запах друг друга.
Учитывая тот факт, что для обеспечения быстрого роста и регенерации растений дернины, а также для пролива мест мочеиспускания и размыва остатков кала (во избежание засоления дернины) требуется большое количество воды, необходимо должное внимание уделить устройству эффективного дренажа, способного за время (1-2 часа) отсутствия лошади в деннике (на тренировке, на прогулке или соревновании) усвоить избыток влаги. Можно устраивать фундамент денников непосредственно на гравии слоем 15-20 см. Фундамент следует изготовить по типу армированной монолитной железобетонной ленты. Такой тип фундамента называется “плавающим” и обеспечивает равномерную осадку под нагрузкой. С целью обеспечения непромерзания зимой подошвы фундамента, под отмостку шириной 1 м вокруг здания следует уложить пенопласт толщиной 4-5 см (или другой теплоизолирующий материал). Для обеспечения работы дренажа необходимо поднять дренажный слой в помещении над уровнем почвы снаружи здания на 20-25 см и устроить его из известкового щебня для обеспечения раскисления дернины. На всю поверхность известкового щебня в деннике расстилается укрывной материал типа “Лутрасил”, пропускающий воду и не пропускающий частицы почвы (для предотвращения заиливания дренажа) и далее устраивается конструкция дернины (Рис.7).

Рис.7 Схема устройства дернины в деннике.
С целью обеспечения необходимой прочности дернины в качестве её несущей конструкции можно предложить устройство слоя из “эффективного” (щелевого) полуторного кирпича уложенного плотно друг к другу вертикально отверстиями с последующим их заполнением грунтопесчаной смесью. При необходимости такая конструкция может быть легко отремонтирована, так как не является цельной. Далее раскатывается готовый газон на тканой основе или устраивается дернина путем засевания грунтовой смеси толщиной 4-5 см специально подобранными для этого семенами злаковых растений.
Для обеспечения быстрого роста и развития растений дернины следует поддерживать температуру 25-26
0С, предусмотреть требуемый полив, подкашивание, хорошую освещенность, и облучение ультрафиолетовыми облучателями типа “Флора”. Через 1,5-2 месяца уже возможно заселение денников лошадьми.
Отмечая положительное влияние ионизации воздуха люстрами Чижевского, академик РАМН В.М. Боголюбов писал: “Воздействие аэроионов сопровождается снижением давления крови, увеличением числа эритроцитов и уменьшением лейкоцитов, ростом гемоглобина. Улучшается общее самочувствие, восстанавливается аппетит и сон. Повышается умственная и физическая работоспособность, ускоряется ход восстановительных процессов в утомленной мышечной системе. Отмечено стимулирующее действие аэроионов на белковый и водный обмены, на синтез витаминов группы В. Стабилизируется уровень кальция и фосфора, снижается содержание сахара в крови. Под влиянием аэроионотерапии повышается устойчивость к различным неблагоприятным факторам, стимулируются защитные силы организма. Показано стимулирующее влияние аэроионов на развитие и рост…”
Известен и тот факт, что применение аэроионизации очень сильно снижает запыленность и содержание вредных микроорганизмов в воздухе помещения, а повышенное осаждение пыли из воздуха на ограждающие конструкции подтверждается личными наблюдениями. Вся взвесь, в том числе и капельная, оседает на стенах и потолке помещения, что хотя и несколько затрудняет их уборку, но весьма значительно улучшает эпизоотическую ситуацию. Вышеизложенное убеждает, что данное свойство аэроионизации следует более настойчиво внедрять при содержании быстроаллюрных лошадей.
К мерам борьбы с запыленностью и загрязненностью микроорганизмами можно отнести такие, как:
- вентилирование помещений;
- применение ультрафиолетовых (УФ) облучателей воздуха;
- более широкого введения в рацион лошадей гранулированных кормов;
- своевременного удаления навоза;
- уборка и дезинфекция конюшен;
- своевременное выявление и изолирование больных животных;
- обеспечение персонала конюшни чистой рабочей одеждой и обувью;
- озеленение окружающей территории.
Солнце – мощный источник инфракрасного и ультрафиолетового излучения. Проникая в организм лошади, лучистая энергия превращается в тепловую, электрическую и химическую. Особенно важным является свойство солнечного света образовывать в организме животных витамин D
3. В качестве искусственных источников ультрафиолетового излучения рекомендуется применять различные лампы ЛЭ, ДРТ, ДБ. В случае применения в конюшне в качестве подстилки дернины, для облучения растительного слоя могут применяться облучатели “Флора”. Желательно в каждой конюшне оборудование соляриев для проведения процедур в холодное время года.
2.2. Технология выращивания жеребят быстроаллюрных пород
В сложной системе выращивания быстроаллюрной лошади, процессы на этапе роста и развития жеребят занимают ведущее положение.
В главе 1 мы уже отмечали значение экологических факторов на воспроизводительную функцию быстроаллюрных лошадей. В комфортных условиях негативное влияние внешней среды минимально. В тоже время отклонения от комфортных условий (сезонные явления, температура, влажность, давление, солнечная радиация, состав пищи, воды, воздуха и др.) могут порождать нежелательные отклонения в развитии эмбриона.
Литературные данные указывают также, что при содержании новорожденных животных в температурных условиях отличных от комфортных могут проявляться:
- неправильное функционирование дыхательной системы;
- нарушение роста и развития;
- недоразвитие мышечной и бурой жировой ткани;
- сдвиг тиреоидного гормонального фона, управляющего термогенезом и программирующего организм к соответствующим фенотипическим проявлениям;
- синдром внезапной смерти новорожденных животных и т.д (11).
Было обнаружено, что причинами синдрома внезапной смерти новорожденных, может служить патология развития терморегуляторной системы, сопряженной с блокадой термогенеза в бурой жировой ткани. Установлено также, что формирование способности регулировать температурный гомеостаз у животных происходит не сразу после рождения, а в течение первых нескольких дней после рождения, называемых молозивным периодом, которые и являются наиболее критическими в формировании терморегуляторной системы (11).
Имеющиеся литературные данные в области этологии лошадей, указывают на то, что воздействием в определенные периоды жизни животного можно вызвать существенные сдвиги метаболических реакций и физиологических функций, а знание особенностей индивидуального развития врожденных форм поведения поможет существенно ускорить и облегчить воспитание, обучение и тренинг молодняка. Учитывая тот факт, что зрелорожденные животных, благодаря хорошо развитой ЦНС, быстро адаптируются в окружающей среде и чутки к слабейшим нарушениям окружающей обстановки, обучение жеребенка, приучение на начальном этапе к тесному контакту с человеком надо начинать с первых дней жизни, пока его реакции не стали консервативными. Период новорожденности у лошадей длится от 1 до 6 часов и эти часы особенно важны для формирования поведения лошади в будущем, а в течение первых 1,5 часов жизни у жеребенка вырабатывается 60 реакций, составляющих половину поведенческого “репертуара” известного на сегодняшний день у лошадей. В этот период жеребята чувствительны к малейшим изменениям окружающей обстановки и тактильным раздражениям в области шеи, туловища и задних ног. На данные раздражения они склонны отвечать увеличенной реакцией, отмечается также тенденция к сильному сопротивлению. Учитывая специфику реакций жеребенка, в литературных источниках предлагается метод удерживания новорожденных жеребят: одной рукой за грудь, другой – за круп, - вырабатывающий навык расслабляться в руках человека по мере повторения такого удерживания. Данное упражнение способствует не только формированию структур нервных связей мозга, но и изменяет биохимию мозга и надпочечников, способствующей адаптации к стрессам. В литературных источниках указывается, что обучение жеребенка следует начинать как можно раньше, с первых часов жизни, пока его поведенческие реакции не обрели консервативность (31). Так оповаживание жеребят, начатое с первых дней жизни, в дальнейшем в основном не вызывает у жеребят стрессорных реакций, сохраняются устойчивые навыки выполнения упражнений, повышается обучаемость. Через неделю, а возможно и раньше, после рождения на жеребенка надевают специальный маленький недоуздок, а при любом выводе кобылы из денника – берут на поводок и ведут рядом с кобылой. Через несколько упражнений жеребенок идет охотно не испытывая опасений и не сопротивляется. Далее можно совершать иные упражнения (чистка, приучение к щетке, прогулки) придерживая жеребенка за недоуздок или поводок, прикрепленный к недоуздку (3). Такие упражнения, начатые с первых часов жизни жеребенка, позволяют перенести в значительной степени у жеребенка реакцию следования на человека, включив его в число биологически важных раздражителей, так как функциональная система следования действует по принципу минимального обеспечения у большинства жеребят не более 4-6 часов после рождения, а второй этап формирования этой системы завершается в основном во вторые сутки жизни. Приводятся также данные о том, что функциональная система прыжка 5-6-месячных жеребят действует по принципу минимального обеспечения в среднем на протяжении первых четырех занятий, а системогенез прыжкового поведенческого акта завершается у жеребят примерно после семи занятий. При этом обучение жеребят происходит в условиях эмоционально-положительного реагирования на преодоление препятствий, в том числе в форме игры. Отмечается также, что навык прыжка у жеребят сохраняется спустя полтора года после раннего обучения, а их нервная и сердечно-сосудистая системы не испытывали перенапряжения во время тренировок. Указывается также, что принудительное напрыгивание, базирующееся на страхе перед человеком и на проявлении пассивнооборонительных реакций, создает неблагоприятные условия для деятельности функциональной системы прыжка. Реакции страха, нарушая координационные связи двигательного анализатора, ухудшают качество прыжка лошади (31). По этим причинам применение знаний этологии является эффективным средством выращивания высококлассных быстроаллюрных лошадей и характеризует данные технологии, как относящиеся к интенсивному типу производства.
Теперь обратимся к соответствующим вышеизложенному материалу моментам традиционным технологиям быстроаллюрного коневодства, реализованных в рысистом, чистокровном верховом и полукровном спортивном направлениях. Так при выращивании рысистых лошадей рекомендуется следующее: “В недельном возрасте жеребенка вместе с матерью выпускают в варок на 20 минут. Время прогулки в варке постепенно увеличивают. Такие прогулки не проводят только в морозную (температура ниже минус 20
0С), ненастную и очень ветреную погоду, чтобы не переохладить и не простудить жеребят… В весеннее-летний период при теплой и сухой погоде они могут гулять несколько часов (2-стр.128)”, а в одном из учебных пособии по “Коневодству” написано следующее: “На 3-4 день после рождения, в зависимости от погодных условий, жеребенка выпускаю на прогулку с матерью (5-стр.109)”. Технология выращивания чистокровных верховых лошадей предусматривает, что, начиная с недельного возраста, на жеребенка надевают недоуздок и приучают его к прогулкам и ходьбе в поводу. С 5-6-го дня жеребят с матками приучают к моциону и прогулкам в паддоках и левадах. В зависимости от погодных условий определяют длительность прогулок на улице, доводя их до 2 ч в течение первых 2 мес жизни... Весной строго следят за тем, чтобы жеребята долго не лежали на непрогретой земле, очень важно избежать простуды в первые дни закаливания (2-стр.139).
Основу поголовья быстроаллюрных лошадей, используемых человеком в конном спорте, составляют лошади полукровных пород. Технология выращивания и тренинга верховых лошадей полукровных пород, указывает на то, что в подсосный период жеребята должны находиться вместе с матерями на пастбище, в конюшнях или базах, а специальный тренинг с ними начинают проводить только после отъема… Целенаправленный тренинг жеребят следует начинать сразу после отъема…(2-стр.145) Осенью, после отъема и в течение зимы жеребят тренируют только рысью. В сильные морозы и гололед тренировки отменяют (2-стр.146).
В пояснении к вышеизложенному, считаем необходимым отметить следующие важные, на наш взгляд, моменты.
Приспособляемость животных к холоду происходит нервно-рефлекторным путем. Самым первым звеном, реагирующим на холод, является нервная система и лишь потом, реакция распространяется на весь обмен веществ.
В процессе эволюции у лошадей сформировалась универсальная системная реакция организма на холодовый стресс – выброс в кровь адреналина – гормона мозгового вещества надпочечников, которым и принадлежит ведущая роль в адаптации (акклиматизации) организма к неблагоприятным факторам внешней среды. После выброса адреналина сужаются периферические сосуды тела – кожи и подкожной клетчатки, кишечника, слизистых оболочек, легочных альвеол и расширяются сосуды мозга, сердца, почек и скелетных мышц (12). Уменьшается легочная вентиляция (снижается глубина и частота дыхания), снижается температура кожи, активизируется патогенная микрофлора. Этот механизм лежит в основе простудных заболеваний с развитием воспаления в органах дыхания, мочевыделения и половой сферы (13). Кровь отходит от периферических участков к “ядру” тела, перераспределяя тепло к более важным жизненным органам и устанавливает помеху проникновения холода в
“ядро” тела. Свойство адреналина “централизовывать” термогенез выработано эволюцией и обусловлено тем, что разные ткани организма реагируют на адреналин по-разному. Так в коже находятся преимущественно a-адренорецепторы, при действии на которых адреналином сужаются сосуды кожного покрова. В то же время в скелетных мышцах находятся в основном b-адренорецепторы, при действии на которых адреналином происходит расширение сосудов. Таким образом, возбуждение a-адренорецепторов приводит к катаболическим реакциям, то возбуждение b-адренорецепторов ведет к ускорению анаболизма (12).
В результате воздействия холода на организм лошади в жировой ткани, в результате окисления жирных кислот, меньшее количество запасается в виде АТФ (“главного энергетика” мышечной деятельности) и большее количество рассеивается в виде тепла, повышая теплопродукцию организма. Из крови жирные кислоты попадают в печень и мышцы, где воздействуют на процесс распада гликогена, как фактор разобщения окисления и фосфорилирования, вызывая меньший конечный выход АТФ и повышая теплопродукцию. Аналогично они “поступают” и с другими жировыми клетками. При частом и длительном воздействии холодового стресса, происходит массированный выброс жирных кислот в кровяное русло, приводящий к увеличению запасов энергетического материала - жира в жировых депо (12). Таким образом, адаптация организма к воздействию холода приводит и к “утяжелению” организма в целом и к “опутыванию” мышечных волокон жировыми депо, механически препятствующих повышению амплитуды и импульса движений лошади.
Следует отдельно оговорить свойства
b-адренорецепторов, активизация которых приводит к усилению анаболических процессов в организме. B-адренорецепторы способны воспринимать гормональные сигналы в виде инсулина, соматотропина и глюкокортикоидов. Нейромедиаторы – дофамин и L-ДОФА, повышающие чувствительность клеток к тестостерону и гормону роста, ещё и активизируя синтез и секрецию этих гормонов, действуют на клетки, в основном, посредством возбуждения b-адренорецепторов (12).
Периодическое воздействие холода на организм лошади приводит к активизации рецепторов ацетилхолина – медиатора, передающего нервный импульс с нерва на мышцу. Активизация выброса ацетилхолина позволяет увеличить теплопродукцию в 4 раза, при этом ни сколько не увеличивая мышечную силу (12). Огромные теплопотери сопровождаются, повышением уровня метаболитов в организме, отвлекая энергетические ресурсы организма на их утилизацию, а также приводят к существенному расходу питательных веществ и, как следствие, к перерасходу
кормов.
При увеличении потребности в энергии клетка в первую очередь использует энергию расщепления АТФ. АТФ – слишком дорогое средство поддержания температурного гомеостаза, поэтому АТФ-зависимые экзотермические процессы замещаются процессами, не связанными с использованием АТФ (15-стр.52).
Холодовый стресс провоцирует резкий выброс в кровяное русло гормонов щитовидной железы – тиреоидных гормонов, обладающих сильным разобщающим действием на окисление и фосфорилирование, и активизирующих термогенез. Кроме того, гормоны щитовидной железы активизируют окисление всех энергетических субстратов, программируя повышенный энергетический фон всего организма в условиях противостояния холоду и проникая в ядро клетки, индуцируют синтез информационных факторов, которые
активизируют синтез белка в митохондриях, которые увеличиваются в размерах и утилизируют почти все энергетические факторы. Таким образом, выброс в кровь тиреоидных гормонов приводит к значительному распаду подкожной жировой ткани на глицерин и жирные кислоты, с выбрасыванием последних в кровь (12).
По мере адаптации организма лошади к холоду происходит активизация обмена веществ в сторону усиления окисления и большим выходом энергии в виде тепла. При этом организм программируется и настраивается на сокращение теплопотерь за счет повышения чувствительности рецепторов к действующим на них веществам.
Взамен активизации
b-адренорецепторов в клетках резко возрастает количество ц-АМФ (циклического аденозинмонофостфат), а активизация b-адренорецепторов нацеливается лишь на запуск фермента аденилатциклазы. Аденилатциклаза, находящаяся в мембране клетки, запускает синтез ц-АМФ, вызывая в клетке комплекс изменений. Адаптация организма к холоду приводит к тому, что вместо запуска длинной цепочки взаимодействий происходит лишь активизация ц-АМФ, вызывающего комплекс анаболических реакций в мышцах, экономя организму много времени и сил (12).
Исходя из вышеизложенного, можно с уверенностью отметить, что приведенные выше извлечения из традиционных технологий быстроаллюрного коневодства не только не способствуют повышению анаболизма и “перекрестной адаптации” с физической нагрузкой, а активным (снижением сократительного анаболизма, повышением изоляционных функций) и пассивным (увеличением жировых депо) образом препятствуют.
Терморегуляционные рефлексы вырабатываются быстро и являются прочными, т.к. играют очень важную роль в поддержании температурного гомеостаза. А их прочность и выраженность зависит от продолжительности и величины перепада температур
(15).
Невозможность проведения тренировок в холодные периоды постоянно нарушает ритмичность тренинга и влечет к снижению суммарного эффекта морфологического оформления адаптации жеребят к физическим нагрузкам. Для осуществления контроля за физической нагрузкой на жеребят, необходимо составлять график и план нагрузок, в которые собираются все виды нагрузок. Энергозатраты по суммированным в графике и плане нагрузкам должны полностью соответствовать энергии, поступающей с кормами.
Приведем для примера ещё несколько спорных, на наш взгляд, рекомендаций по тренингу и содержанию жеребят.
“
Навоз из денников убирают ежедневно утром и вечером, а из помещений для группового содержания – по мере надобности (обычно 1-2 раза за стойловый период); подстилку обновляют ежедневно (4-стр.121)”.
“Моцион жеребят, способствующий развитию и укреплению костяка, сухожильно-связочного аппарата, сердечно-сосудистой и мышечной систем, легких, не должен прекращаться с окончанием пастбищного сезона. Прежде всего, отъемышам предоставляются огороженные площадки около конюшни – варки для пребывания на свежем воздухе возможно дольше. Варки целесообразно располагать на достаточно высоком месте, чтобы жеребята могли находиться в них и в сырую погоду (2-стр.131)”.
“После отъема от матерей жеребята как можно дольше должны находиться на свежем воздухе (4-стр.123)”.
“Наряду с содержанием в варках отъемышей начинают тренировать. Групповой тренинг вырабатывает у жеребят производительные движения и выносливость, подготавливает к индивидуальному тренингу и ипподромным испытаниям (2-стр.131)”.
“Жеребят тренируют 6 дней в неделю. Перед групповым тренингом они должны находиться в варках не менее 1ч. В первую неделю тренинга жеребята пробегают и проходят дистанцию 2-3 км. Сначала они свободно пробегают любым аллюром 600-800 м. Следующие 600-800 м преодолевают спокойной рысью, и заканчивается дистанция шагом (2-стр.132)”.
Технология выращивания чистокровных верховых лошадей предусматривает, что до 12-месячного возраста жеребят разбивают по полу, а при дальнейшем тренинге увеличивают нагрузки, вводя два реприза рыси и один реприз кентера. Общий объем тренинга -5-7 км продолжительностью 40 мин. После заездки полуторагодовалых жеребят начинают тренировать в группе. Первую неделю ограничиваются 5-6 мин езды рысью и не менее 25 проводки шагом (2-стр.140-141).
“В течение зимы дистанцию тренировки постепенно увеличивают и к весне доводят до 10-11 км… Протяженность дистанции и скорость её прохождения могут быть уменьшены в связи с состоянием жеребят и погодными условиями (2-стр.132)”.
Технология выращивания и тренинга верховых лошадей полукровных пород указывает, что в специализированные тренировочные отделения конных заводов для спортивного тренинга отбирают молодняк в возрасте 1,5-2 лет из табунов (2-стр.154).
“Обращают внимание на рост, развитие, экстерьер лошадей. В первую очередь ставят в тренинг более рослых, развитых животных (2-стр.154)…”.
На наш взгляд, все перечисленные выше рекомендации имеют ряд существенных изъянов, препятствующих выращиванию высококлассных быстроаллюрных лошадей:
- Скорость прохождения дистанции (степень эксплуатации) определяет уровень теплопродукции и режим теплообмена с окружающей средой. Данному уровню теплообмена должны соответствовать оптимальные условия внешней среды, а каждому режиму нагрузки должен соответствовать и практически применяться определенный температурно-влажностный режим. Этот процесс должен обязательно регулироваться человеком, и вероятно, что эффективным средством достижения этой цели станет применение тренировочной одежды и регуляция микроклимата в специализированных тренировочных помещениях. В настоящее время рынок предлагает очень незначительный выбор тренировочной экипировки для лошадей, которая, к тому же, лишена возможности регулировать уровень теплоотдачи. Для этой цели можно разработать модели тренировочных накидок со сбрасываемыми элементами. Количеством слоев сбрасываемых элементов можно было бы регулировать уровень теплоотдачи, начиная с наиболее нагреваемых частей тела лошади с группами крупных мышц (шея, холка, лопатка, поясница, круп) и далее с менее нагреваемых (плечо, грудная клетка, голень, грудина). Для фиксации сбрасываемых элементов можно применять быстро отстегиваемые крепления. В отношении мышц спины, также необходимы специальные решения, вероятно с применением клапанов для быстрого теплоотвода, а также современных синтетических материалов применяемых для охлаждающих накидок и попон.
- В тренинг попадает молодняк с уже оформившимся в интерьере лошадей привычным уровнем физических нагрузок, определенной мышечной массой и соотношением двигательных единиц (мышечных волокон) с различным типом обмена. По этим причинам индивидуальный тренинг лошадей следует начинать, как можно раньше. Он должен быть научно обоснованным и методически грамотно нацеленным на достижение лошадью навыков определенной специализации.
- Тренинг должен удовлетворять критериям постепенности и равномерности нагрузки, т.е. нагрузка не должна быть “рваной”, должна быть без длительных перерывов во времени. При резком перерыве физической нагрузки, организм лошади начинает резко адаптироваться к новым к условиям, отвечая на эти условия морфо-физиологической реакцией.
- Велико пренебрежение влиянием экологических факторов внешней среды и микроклимата конюшен, что характеризует традиционные технологии в быстроаллюрном коневодстве России, как технологии экстенсивного типа. Они отличаются высокой экономичностью и дешевизной выращиваемых лошадей посредственных потребительских достоинств.
- В тренинг попадает молодняк, благополучно переживший табунные условия содержания, а непереживший эти условия молодняк, по различным условиям (падеж, отставание в развитии из-за частых заболеваний и т.п.) выбраковывается, а вместе с этим и выбраковывается наиболее ценный генетический совершенный для целей отбора по резвости племенной материал.
2.3.
Эксплуатация, тренинг быстроаллюрной лошади и организация моциона племенных быстроаллюрных лошадей
Касаясь вопросов эксплуатации и тренинга, следует особым образом выделить влияние экологических факторов на хозяйственно-полезные качества быстроаллюрной лошади.
В п.2.1. мы уже описывали возможные последствия от воздействия на организм лошади аммиака образующегося при разложении органических азотсодержащих соединений под действием уреазоактивных анаэробных бактерий и сероводорода, образующегося при гниении серосодержащих белковых соединений. Помимо прямого вредного воздействия аммиака и сероводорода имеет место и отвлечение энергетических ресурсов организма (прежде всего, АТФ) на утилизацию этих вредных газов. Тем не менее, как и прежде: “Спортивную лошадь содержат в деннике, в светлой, сухой и хорошо вентилируемой конюшне. В качестве подстилки можно применять опилки и солому. За чистотой подстилки в деннике необходимо постоянно следить (2-стр.169)”. Данный подход является пассивным средством борьбы за чистый воздух в конюшнях, таким образом, мы боремся не причиной образования вредных газов, а с последствиями разложения мокрой подстилки, а на реализацию активных способов борьбы с причинами появления вредных газов у нас не хватает решимости. Тем не менее, именно активные способы помогли бы нам кардинально изменить ситуацию в содержании лошадей, не говоря уже про эстетические моменты от реализации такого подхода к данной проблеме.
Уход за подстилкой сопряжен с субъективной оценкой её состояния, её влажности, а влага в соломе и опилках присутствует всегда. Таким образом, образование из источника возникновения вредных газов полностью предотвратить невозможно, а можно лишь уменьшить их выделение.
В предыдущих параграфах мы уже обращали внимание на влияние адаптационных процессов к длительному воздействию холода на организм животного. Практически вся территория России имеет климатические условия со значительными перепадами зимних и летних температур. Имеются литературные данные, указывающие на то, что формирование химической терморегуляции происходит лучше в условиях значительных перепадов температур среды (22-стр.226), а именно в умеренном и континентальном климате Евразии. Поэтому любые рекомендации по способам выдержки и тренинга быстроаллюрных лошадей с применением или упоминанием холодовых технологий должны восприниматься нами критически. Так, например, имеется следующая рекомендация: “Для содержания кобыл с жеребятами, жеребцов и молодняка всех возрастов, а также в манежах для тренинга лошадей температура воздуха должна быть 16оС, но не менее 4оС (!)…” (2-стр.256). Подобные рекомендации вызывают двойную критическую волну не только потому, что допускают значительное понижение температуры среды при выдержке и тренинге лошадей, но ещё и потому, что не разделяют по условиям среды помещения для содержания и тренинга лошадей при значительно отличающейся у них интенсивности обменных процессов. Различные уровни метаболических процессов требуют наличия соответствующих и различных, в первую очередь, температурно-влажностных условий пребывания в них лошадей. Под этими условиями мы понимаем комфортные и оптимальные условия среды. О комфортных условиях мы говорим, когда лошадь отдыхает (полный покой). Данные условия характеризуются самым низким энергетическим обменом (36-стр.25), а количество теряемого тепла равно количеству вырабатываемого тепла (36-стр.40). Температурный интервал, характерный для комфортных условий носит название зоны термонейтральности, при котором обмен веществ поддерживается на постоянном минимальном уровне (36-стр.71). При этом мы понимаем, что всякая деятельность (мышечная, пищеварение) приводит к перегреванию животного, что приводит к активации системы термогенеза. Об оптимальных условиях мы говорим в период организации моциона, тренинга и эксплуатации, поскольку в эти периоды на лошадь оказывает воздействие физическая нагрузка и о комфорте речи идти уже не может. Кроме того, мы подразумеваем, что при оптимальных условиях эксплуатации и тренинга достигается наивысший КПД мышечной деятельности (для каждого уровня интенсивности эксплуатации), при наименьших затратах энергии и минимальном уровне потребления кислорода.
Так бытует мнение и не только бытует, но широко распространено в практическом использовании, что в зимнее время в конюшне температура воздуха не должна превышать 10-12оС, т.е. когда лошади ещё не дрожат, но находятся при повышенном тонусе. Считается также, что повышенная температура, при недостатке внимания к лошадям, делает их вялыми и ленивыми. Обратимся за разъяснением сути происходящих при т.н. “пограничной” температуре 10-12оС в организмах лошадей термоэнергетических процессов к литературным источникам.
Установлено, что на периферии (кожа и подкожные сосуды) холодовые рецепторы проявляют максимальную активность в диапазоне 16-32оС, они дают фазовое учащение разряда на понижение температуры и количественно преобладают над тепловыми (24-стр.56).
Так ещё в 19 веке было обнаружено, что химическая терморегуляция может проявляться и при отсутствии видимой сократительной деятельности мышц. При этом было замечено то, что холодовая мышечная дрожь у акклиматизированных к холоду животных выражается значительно слабее, нежели у неакклиматизированных и делается вывод о том, что при адаптации к холоду сократительный термогенез мышц (холодовая дрожь) уступает место несократительному термогенезу, который в последствии был назван, как терморегуляционный тонус, который является специфической реакцией организма в ответ на охлаждение. Исследования показали, что при интенсивном терморегуляционном тонусе обмен в мышцах у мышей повышался на 120-155% от исходного уровня, при этом делается вывод о том, что при высоком терморегуляционном тонусе возможно повышение обмена на 36-45% и эти цифры почти совпадают с цифрами общего газообмена. Вместе с тем, холодовая дрожь, остается более эффективным энергетическим механизмом, т.к. потребление мышцами кислорода может повышаться до 400% (23).
Обнаружено, что при терморегуляционном тонусе происходит микровибрация (холодовый тремор) – асинхронные сокращения отдельных мышечных волокон или небольших групп в определенной последовательности, при этом общее тоническое укорочение мышц весьма незначительное и заметно только при постановке специального опыта. Это явление и составляет сущность терморегуляционного тонуса мышц и является причиной микровибрации мышц организма при полном видимом покое, в то время как при холодовой дрожи сокращается большая масса мышечных волокон, при которой наблюдается некоторое постоянное тоническое сокращение мышц. Тремор сохраняется и после наступления холодовой дрожи и как бы заполняет паузы между приступами дрожи. Эффективность холодовой мышечной дрожи, с точки зрения терморегуляции, в 2,5 раза выше мышечной работы и является мощным механизмом химической терморегуляции. Вместе с тем, наиболее выгодным источником теплопродукции является терморегуляционный тонус по ряду причин:
- приток крови к мышцам и теплоотдача с мышц при треморе меньше чем при дрожи,
- терморегуляционный тонус более строго локализован топографически в проксимальных мышцах тела (шея, голова), что позволяет избегать дополнительных потерь тепла (23), хотя более детальные исследования показали, что холодовый тремор присущ практически всем мышцам тела за исключением мышц диафрагмы (24-стр.44).
У зрелорождающихся животных специфические виды мышечной деятельности (тремор, холодовая дрожь) становятся сформировавшимися уже с первых дней и даже часов своей жизни, ведь терморегуляционные рефлексы вырабатываются быстро и являются прочными.
Значение терморегуляционного тонуса для целей выживания организма очень велико, т.к. ещё до наступления холодовой дрожи, такая скрытая сократительная деятельность мышц обеспечивает достаточно высокий объем химической терморегуляции.
Вместе с тем, необходимо отметить, что как холодовый тремор, так и холодовая дрожь, как проявления деятельного состояния организма при адаптации к холоду, сопровождаются уменьшением к.п.д. мышечного сокращения (в
1,5-3 раза!!!), т.е. на любую единицу мышечной работы тепла производится больше, чем в норме. Одновременно, происходят те же морфо-физиологические процессы в мышечной ткани и органах, которые мы уже описывали в параграфах 2.1 и 2.2.
По этим причинам, умиление коневодов быстроаллюрных лошадей состоянием своих подопечных в фазе интенсивного тремора – необоснованно с точки зрения зоотехнии и неоправданно с точки зрения цели специализации таких лошадей. Именно в результате интенсивного терморегуляционного тонуса происходит постоянное понижение основных полезных качеств животных, “имущество” как бы самоуничтожается невидимо глазу и становится малоценным для целей разведения при полном попустительстве и даже удовольствии коневодов видимой частью этого явления.
Любой организм, проявляя ту или иную двигательную активность, постоянно приспосабливается в использовании своих морфофункциональных резервов. При этом двигательная активность может существенным образом влиять на состояние организма, как в сторону увеличения функциональных и структурных резервов, так и в сторону их уменьшения.
Спортивная тренировка является адаптационным процессом, направленным на приспособление организма к изменениям химизма мышц, органов и внутренней среды при осуществлении мышечной деятельности, причем физическая нагрузка для получения устойчивого тренировочного эффекта должна оказывать стрессорное воздействие и быть стабильной. Кроме того, при постоянной тренировке увеличивается масса мышц, в них повышается количество сократительных белков, растет содержание гликогена, АТФ и креатинфосфата, возрастает сила и скорость мышечного сокращения, развивается полезная гипертрофия (
18-стр.31).
Однако немаловажен и характер тренировочного процесса. Так в отличие от интервального типа тренировки, при длительных нагрузках имеющих, хотя и высокий, но не максимальный уровень, развиваются процессы схожие с длительным действием холодового фактора, а именно:
- увеличивается доля красных (медленных) и промежуточных мышечных волокон, уменьшается доля белых (быстрых) волокон,
- увеличивается доля неэстерифицированных жирных кислот и глицерина по сравнению с долей содержания глюкозы и лактата, что говорит о преимущественном вкладе липидной составляющей в энергетику мышечного сокращения при адаптации к длительным нагрузкам,
- достоверное изменение спектра жирных кислот общих липидов крови (снижение доли содержания короткоцепочечных жирных кислот и повышение коэффициента ненасыщенности) (21-стр.71).
В процессе работы животного начинает развиваться мышечное утомление, вызванное накоплением продуктов обмена веществ (в частности, лактат, образующийся при расщеплении гликогена), угнетающие сократительную деятельность мышц. Также постепенно истощаются и энергетические запасы в отдельных органах и тканях, происходит резкое уменьшение запасов гликогена, нарушаются процессы ресинтеза АТФ и креатинфосфата. Кроме того, мышечные волокна не могут работать при любой температуре без денатурации, поэтому ограничения, налагаемые на деятельный диапазон температур зависит и от степени энергообразования в двигательных единицах, что также может влиять на степень угнетения мышечной деятельности. Однако случается, что утомленное животное после изнурительной работы, крупными шагами, или даже легкой рысью бежит к табуну или возвращается в стойло. Это обстоятельство означает, что клетки коры больших полушарий, благодаря эмоциональному возбуждению, влияют через подкорковые образования и стволовую часть головного мозга на метаболические процессы в утомленных органах и тканях (18-стр.20). Для обеспечения более эффективного тренинга и восстановления после него, следует шире внедрять технологии, основанные, как на знании этологии, так и степени тренированности лошадей.
Приведенные факты говорят о необходимости методичного внедрения в быстроаллюрном коневодстве интервального тренинга, основанном на адаптации организма лошади к кратковременным нагрузкам максимальной интенсивности, без развития значительной степени утомления.
Для достижения наибольшего уровня тренированности скаковой (рысистой) лошади, необходимо проводить целенаправленную работу на корректировку в мышцах лошадей двигательных единиц с гликолитическим, оксидативным и оксидативно-гликолитическим типом обмена в зависимости от уже достигнутого уровня тренированности и длины основной дистанции, на которую предполагается лошадь тренировать. Можно уверенно предположить, что достижение этой цели прямо зависит от назначения определенного количества (3…5) по длительности (200…400…700м) интервалов репризов с максимальной резвостью. Таким образом, можно повышать у лошади, обладающей спринтерскими задатками выносливость к пролонгированным нагрузкам, так и у лошади, обладающей стайерскими способностями, повышать резвость. Это обстоятельство выдвигает методики, основанные на адаптации лошадей к различным режимам коротких максимальных физических нагрузок, в категорию наиболее эффективного инструментария для тренировки быстроаллюрных лошадей.
Учитывая, что основные традиционные призы на ипподромах разыгрываются в интервале от 1000 до 3200 м, для половозрастных групп 2, 3 и 4 лет, мы можем рекомендовать строить тактику интервального тренинга с учетом выбора основной дистанции в скаковом (беговом) сезоне на которую планируется записать лошадь, учитывая индивидуальные особенности животного. При этом учитывается то обстоятельство, что длительность резвого приема не должна быть дольше 30 с (37). Это обеспечивает преимущественное развитие анаэробного процесса энергообеспечения перед аэробным.
Так вклад анаэробного и аэробного процессов энергообеспечения при выполнении 3-х минутных интенсивных физических упражнений, составляет (в%): 80/20 при работе в течение первых 30 с; 45/55 – от 60-й до 90-й с; 30/70 – от 120-й до 180 с (37 – стр.10).
Двигательная гипоксия при напряженной мышечной деятельности сокращается по мере адаптации к физической нагрузке, возрастает мышечная масса и совершенствуется система транспорта кислорода к тканям, что означает повышение уровня тренированности лошадей. Установлено, что изменения биохимических показателей при однократной нагрузке свидетельствовали о торможении гликолитических процессов. В то же время при интервальной нагрузке обнаружено повышение активности гликолиза (
25).
Назначение режимов максимальной нагрузки и длительности интервалов отдыха между ними, производится с учетом индивидуальных особенностей лошадей и момента наступления суперкомпенсации (26) после первого и последующих резвых приемов, о которой можно судит по активности альдолазы, степени насыщения венозной крови кислородом и содержанию фосфора. В качестве примера, можно рекомендовать следующий алгоритм подготовки 2-летней скаковой лошади на дистанцию 1000 метров. Данная дистанция делится на 4 “резвых” интервала (200+250+250+300). После первого резвого галопа (300м) переходят на кентер (300м), потом рысь (500м) и шаг (4 минуты), далее резвый галоп (250м), потом на рысь(300-500м) и шаг. Через 15 минут вновь поднимают на кентер (150-200м) и переходят на резвый галоп (250м), потом переходят на кентер (500-700м), на рысь(300-500м) и шаг. Через 30 минут вновь поднимают на кентер (150-200м) и переходят на резвый галоп (200м), потом переходят на рысь(300-500м) и лошадь “вышагивают”. При этом очень важно, чтобы резвый галоп проводился при максимальной нагрузке (25). Учитывая степень тренированности, интервалы отдыха между резвыми приемами следует сокращать, доведя их до 8-10 (после второго резвого приема) и 12-15 минут (после третьего резвого приема).
Основным недостатком интервального метода тренировки, рекомендованного ВНИИК, по нашему мнению, остается пренебрежение различиями процессов теплообразования и соответствующего ему теплообмена организма со средой в режимах резвого галопа, кентера, рыси и шага, по причине того, что каждому из этих режимов соответствуют определенно различные оптимальные условия среды и, в первую очередь, температурно-влажностные.
Другим существенным недостатком метода интервального тренинга составляют значительные временные перерывы (несколько дней) между днями проведения интервального тренинга. Данный режим ограничивается периодами полного восстановления мышечной ткани и органов после максимально интенсивной нагрузки. Тем не менее, опыт конного завода “Восход” показал, что интервальные нагрузки можно проводить в 2 раза чаще с интервалом в 4-5 дней (27). Но и это вряд ли можно считать пределом. При условии широкого применения в скаковом коневодстве средств ускорения восстановления функционального состояния лошадей после интенсивных нагрузок:
- кислородная терапия в первые минуты после нагрузки,
- массажные процедуры,
- ультрафиолетовое и инфракрасное облучение,
- применение биологически активных добавок (янтарная кислота, глутаминовая кислота, натриевая соль АТФ, оротат калия, карбоксилаза, фолиевая кислота, пангамат кальция),
- применение специальных подкормок,
- водный раствор мелассы, вспененный газообразным кислородом (28), - перерывы между днями интервального тренинга можно сократить ещё. Для достижения наибольшего эффекта интервального метода тренинга может стать применение комфортных условий содержания, оптимальных условий моциона, эксплуатации и тренинга (организация микроклимата в специальных помещениях для тренинга, специализированной тренировочной одежды), адекватного сбалансированного кормления, что пока не имеет практического применения, как на территории России, так и за рубежом, что подтверждается отсутствием каких-либо литературных данных по данному кругу вопросов. В настоящее время рынок предлагает коневодам очень узкий ряд моделей тренировочных накидок (Рис.8-9), который, вместе с тем, быстро развивается:

Рис.8 Европейская водонепроницаемая тренировочная накидка.
В дождливую погоду её можно постелить поверх седла.

Рис.9 Пропускающая тренировочная попона. Сохранит лошадей теплыми и сухими во время тренинга.
Центральной реакцией начального теплообразования является гидролиз АТФ миозиновой АТФ-азой в момент мышечного сокращения. Теплообразование, возникающее после мышечного сокращения, связано с окислительными процессами ресинтеза АТФ. Поэтому возрастание потребления кислорода в организме при мышечной работе (в том числе и во время холодовой дрожи) отражает возрастание окислительных процессов ресинтеза макроэргов и используется для оценки так называемой химической терморегуляции. Во время холодовой дрожи потребление кислорода у гомойотермных организмов возрастает в 3-9 раз. Учитывая тот факт, что у адаптированных к холоду животных теплопродукция мышечного сокращения за счет механизма химической терморегуляции возрастает до 400%, адаптация к холоду имеет обратную корреляцию по обеспечению кислородом тканей и органов к уровню работоспособности лошади.
Бытует мнение, что мускулатура быстроаллюрных лошадей, селекционируемых столетиями на резвость представлена быстросокращающимися волокнами (29), однако имеются литературные данные, свидетельствующие о том, что даже у лошадей чистокровной верховой породы в средней ягодичной мышце могут отсутствовать полностью или содержаться менее 30% двигательных единиц с чисто гликолитическим типом обмена, т.е. быстрых волокон (30). Кроме того, имеются данные при изучении влияния кратковременных высокоинтенсивных тренировок (4…6-недельная тренировка в велосипедном спринте) на структуру скелетных мышц о том, что происходило снижение содержания красных (медленных) волокон с 57% до 48% и увеличении белых (быстрых) волокон с 32% до 38% (37-стр.32). Данные обстоятельства лишний раз подтверждает значение научно-обоснованного подхода в применении тех или иных методик для достижения целей специализации лошадей. Так во многом увеличение длительности тренировочного процесса без максимальной нагрузки схоже по своему морфо-физиологическому эффекту с длительным воздействием холода на организм животного, так как оказывает прямое действие на мышечную структуру, увеличивая долю волокон с оксидативным и гликооксидативным типами обмена, а также формирует структуру энергетических депо.
Существенным резервом повышения качества выращиваемого поголовья быстроаллюрных лошадей на конных заводах России, может стать широкое применение модельных интерьерных характеристик лошадей на разных стадиях роста, тренинга, эксплуатации и оценки племенной ценности. Так при помощи:
- иммуногистохимических методов можно определить структурный состав (быстрые, медленные и промежуточные волокна) мышечной ткани,
- биоимпедансного анализа осуществить динамический контроль состава тела (жировой ткани, безжировой ткани, мышечной массы, общей воды в организме), динамический контроль водных секторов организма – клеточной, внеклеточной и интерстициальной жидкостей и т.д. (32),
- биохимического анализа крови определяют кислородно-транспортную функцию крови (гемоглобин, эритроциты, гематокрит), активность анаэробного процесса (активность альдолазы, содержание лактата, насыщение венозной крови кислородом, содержание фосфора), активность аэробного процесса (содержание пирувата, активность каталазы), активность белкового обмена, содержание различных форм кальция, фосфора и магния (29),(33),(34),
-
использование возможностей термовидения, при осуществлении контроля температуры тела лошадей для определения уровня их тренированности и корректировки тренировочных методик в целях повышения работоспособности (Рис.10-13) (39-стр.104).

Рис.10 Термограмма лошади, перенесшую физическую нагрузку максимальной мощности.

Рис.11 У данной лошади температура задней части тела выше, чем передней

Рис.12 Термограмма лошади, показавшей лучший результат на скачках. Температура равномерно распределена по всему телу и при этом наивысшая

Рис.13 Термограмма лошади выполняющую легкую физическую работу.
Однако вывод этого литературного источника о том, что система термовидения позволит когда-то предсказать победителя скачек выглядит пока неубедительным, т.к. температура тела лошади означает, прежде всего, уровень теплообразования, в том числе и от процесса химической терморегуляции. По этой причине данная методика может эффективно использоваться для контроля за уровнем тренированности лошади, а точность оценок – от условия применения других методов контроля за тренированностью лошадей.
Модельные интерьерные характеристики и данные методы анализа и контроля должны представлять явный интерес для специалистов-коневодов по причине возможности эффективного применения данных методов и относятся, вместе с методами ускорения восстановления спортивных лошадей после интенсивных физических
нагрузок, к интенсивному типу технологий. Однако при применении комфортных технологий конкретные контрольные цифры, полученные на лошадях, выращенных при сильном влиянии экстенсивных технологий, могут приниматься лишь к сведению, по причине различия у этих лошадей (выращенных в интенсивных и экстенсивных технологиях) обменных процессов.
(Продолжение следует)
Литература
- Щеглов Е.В., Попов В.В. Разведение сельскохозяйственных животных. – М.: КолосС, 2004. – 120 с.
- Калашников В.В. и др. Практическое коневодство. Справочник. – М.: Колос, 2000. – 376 с.
- Парфенов В.А. Лошади. – М.: Народное творчество, 2002. – 192 с.
- Федотов П.А. Коневодство. – М.: Колос, 1981. –240 с.
- Красников А.С., Хотов В.Х. – Коневодство. – М.: Изд-во МСХА, 1994. – 192 с.
Miodrag Dodic, Arianne Peers, John P. Coghlan, Marelyn Wintour — Trends in Endocrinology and Metabolism 1999, 10:86-91
http://www.cironline.ru/articles/pregnancy/25/
Рыжавский Б.Я. Развитие головного мозга в ранние периоды онтогенеза: последствия некоторых воздействий, ДГМУ, 2000, Биология,
http://www.pereplet.ru/obrazovanie/stsoros/922.html
Елисеенко О. Журнал "Мир Скачек" №4 – 2002
http://www.dark-horse.narod.ru/
Кузнецов А.Ф. и др. Гигиена животных. – М.: Колос, 2001. – 368 с.
Лазарев Н.В., Гадаскина И.Д. Вредные вещества в промышленности. Справочник. Том III. – Л.: Химия, 1977. – 608 с.
Автореферат диссертации. Исследование структурно-функциональных основ динамики реакций новорожденных в ответ на действие внешних раздражителей в процессе постнатального онтогенеза.
Температурные стимуляторы анаболизма Ю.Б. Буланова.
http://athlete.ru/?part=pharmacology&txt=temp_bul
Волков Л.Б. Физиологические основы терморегуляции. – Харьков, 2003.-
http://rukzachok.com.ua/
Пугачев М.К. и др. Морфофункциональные изменения коры надпочечников крыс при общем
внешнем перегревании в сочетании с наркозом. – Смоленск: СГМА, 1997. – 4с.
15. Хаскин В.В. Энергетика теплообразования и адаптация к холоду. - М.: Наука, 1975. – 200 с.
- Алексеев Н.Д. Адаптация лошадей к температурным факторам среды. Диссертация на
соискание ученой степени кандидата биологических наук. – ВНИИК, 1985. -199с.
- Иванов К.П. О биоэнергетических механизмах гомойотермии. Журнал общей биологии. Том
51. №1 – М.: 1990. – стр. 36 -53.
- Лазарев В.М. Физиология движения и продуктивность сельскохозяйственных животных.
Лекция. – Саратов, 1985. – 60с.
- Якименко М.А. О приросте тепла в сокращающейся скелетной мышце при адаптации к
холоду. Сборник трудов под ред. Иванова К.П. и Слонима А.Д. – Новосибирск.: Наука. Сиб.
отд., 1975. – 13с.
- Филипченко Р.Е. Активность сукцинатдегидрогеназы в различных группах скелетных мышц
белых крыс в норме и после холодовой адаптации. Сборник трудов под ред. Иванова К.П. и
Слонима А.Д. – Новосибирск.: Наука. Сиб. отд., 1975. – 14-17с.
- Волкова Е.В. и др. Исследование энергетического метаболизма при длительных физических
нагрузках. Всесоюзный симпозиум. Сборник тезисов. – Тбилиси.: Мецние реба, 1987. – 71с.
Слоним А.Д. Химическая терморегуляция в онтогенезе и у взрослого животного. Успехи
современной биологии. Том 47. Вып.2. – Ленинград. – 220-231с.
23. Иванов К.П. Мышечная система и химическая терморегуляция. – М.-Л.: Наука, 1965. – 119с.
- Лупандин Ю.В. Двигательные механизмы защиты организма от холода. – Л.: ЛГУ, 1988. –
173с.
Ласков А.А. Рекомендации по интервальной тренировке скаковых лошадей. – ВНИИК,1979.
– 10с.
Сергиенко Г.Ф. Физиологические и биохимические аспекты тренинга быстроаллюрных
лошадей. – ВНИИК, 1997. – 34с.
Леонова М.А., Шишкин Е.М. Опыт ипподромного тренинга скаковых лошадей конного завода
“Восход”. – ВНИИК, 1984. – 30с.
Ласков А.А. и др. Средства ускорения восстановления функционального состояния
спортивных лошадей после интенсивных физических нагрузок. – ВНИИК, 1988. – 22с.
Сергиенко Г.Ф. Биохимические основы тренировки верховых полукровных лошадей. –
ВНИИК, 1984. – 76с.
Сноу Д. и др. Состав мышечных волокон и истощение запасов гликогена у лошадей при
испытании на выносливость в ходе марафонской скачки. Перевод с английского. – М.:
ВНИИК, П-418, 1982.-15с.
Лытнева Е.В. Этологические аспекты выращивания спортивной лошади. – ВНИИК, 1987. –
118с.
Матыцин О.В. и др. Спортивная медицина и исследования адаптации к физическим
нагрузкам. Научные чтения. – М.: Типография Россельхозакадемии, 2005.- 282с.
Сергиенко Г.Ф., Сергиенко С.С. Оценка уровня общей тренированности верховых лошадей,
выступающих в соревнованиях по конному спорту. – Дивово: ВНИИК, 1996. – 48с.
Ласков А.А. Подготовка лошадей к олимпийским видам конного спорта. – ВНИИК, 1997. –
244с.
Тепловая адаптация (акклиматизация). – Москва: Физкультура и спорт, 2004.-
http://sportlib.ru/books/reabil/fiziol/glava28.htm
Степанов Д.В., Родина Н.Д. Экологический подбор в животноводстве. – Орел: ОГУ, 2004. –
408с.
Лоуренс Спрайет, Марк Харгривз и др. Метаболизм в процессе физической деятельности. –
Киев: Олимпийская литература, 1998. – 288с.
38. Зайцев С.Ю., Конопатов Ю.В. Биохимия животных. Учебник. – СПб: Лань. 2004 – 384с.
39. “Конный мир”. Журнал. №1. 2003г.

Используются технологии
uCoz